

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Thyroid hormones regulate developmental processes such as neurogenesis, myelination, dendrite proliferation and synapse formation (Bernal et al., 2003; Williams, 2008). In particular, maternally synthesized thyroid hormones at very late embryonic stages influence neuronal proliferation and migration of neurons in the cerebral cortex, hippocampus and medial ganglionic eminence (Narayanan & Narayanan, 1985; Ausó et al., 2004; Cuevas et al., 2005). In addition, a close association exists between thyroid hormones and brain cholinergic function (Smith et al., 2002). These effects are mainly observed in specific cholinergic nuclei and their pathways, such as the basal forebrain and the hippocampus (Patel et al., 1987).
Hippocampal neurons are vulnerable to diabetes (Gispen & Biessels, 2000; Magariños & McEwen, 2000); memory loss and impaired executive function also accompany type 2 diabetes (Ryan & Geckle, 2000). In addition, diabetes reduces neuroblasts in the dentate gyrus of the hippocampus in type 1 (Jackson-Guilford et al., 2000; Beauquis et al., 2006) and type 2 (Hwang et al., 2008) models; these neuroblasts extend their axons and contact CA3 pyramidal neurons in the hippocampus proper, becoming integrated into the hippocampal circuitry (Stanfield & Trice, 1988; Hastings & Gould, 1999). In diabetic rats, hyper-activation of the hypothalamo-pituitary adrenal axis is well described, and corticosterone in the adrenal gland mediates diabetes-induced impairments of hippocampal synaptic plasticity and neurogenesis, as well as associated cognitive deficits (Landfield et al., 1978; Trudeau et al., 2004; Montaron et al., 2006; Stranahan et al., 2008). The correlation between thyroid hormones and adrenal corticosteroids hormones has been reported (Silva & Bianco, 2008).
Although there are reports about the effects of hypothyroidism in the type 1 (Hibbe et al., 1991) and type 2 diabetic models (Matsushita et al., 2005; Tamura et al., 2005; Hwang et al., 2009), no studies have been reported about the effects of hypothyroidism in type 2 diabetic model on cell proliferation and neuroblast differentiation. In the present study, we investigated the consequences of adult-onset hypothyroidism in diabetic rats using methimazole, an anti-thyroid drug, which has been used in the management of hyperthyroid patients (Cooper 2005). We also investigated how the hypothyroid state modifies neuroblast differentiation which retarded by diabetic state possibly through high corticosterone level in the hippocampal dentate gyrus of Zucker diabetic fatty (fa/fa, ZDF) rats by measuring expression of Ki67, an endogenous marker of proliferation expressed during late G1, S, M and G2 phases of cell cycle (Cooper-Kuhn & Kuhn, 2002), and doublecortin (DCX), a marker of neuronal progenitors differentiating into neurons (Karl et al., 2005).
Materials and Methods
Experimental animals
Male and female Zucker diabetic heterozygote rats (fa/+) were purchased from Genetic Models (Indianapolis, IN, USA) and mated each other. They were housed in a conventional state under adequate temperature (23℃) and humidity (60%) control with a 12-h light/12-h dark cycle, and free access to food and water. Purina 5008 rodent diets (7.5% fat) were provided as recommended by Genetic Models. The procedures for handling and caring for the animals adhered to the guidelines that are in compliance with the current international laws and policies (NIH Guide for the Care and Use of Laboratory Animals, NIH Publication No. 85-23, 1985, revised 1996). All of the experiments were conducted to minimize the number of animals used and the suffering caused by the procedures used in the present study.
Genotyping of fa gene and experimental design
Genotype of fa gene herein was determined with the strategy described previous our study (Hwang et al., 2008, 2009). ZDF rats were randomly divided into 2 groups (n=7 per group) with vehicle-ZDF and hypothyroid-ZDF group. At 7 weeks of age, hypothyroidism was induced by the administration of 0.03% 2-mercapto-1-methyl-imidazole (methimazole, Sigma, St. Louis, MO, USA) in drinking water for 5 weeks. ZLC rats (n=7) were served as the control. All animals were euthanized at 12 weeks of age.
Measurements of levels of blood glucose, serum thyroid hormones, and serum corticosterone
To measure blood glucose concentration, blood was analyzed by using a blood glucose monitor (Ascensia Elite XL Blood Glucose Meter, Bayer, Toronto, ON, Canada). To confirm the hypothyroid state and corticosterone levels, the animals were anesthetized with 60 mg/kg chloral hydrate and blood specimens were drawn from the right ventricle of ZLC, ZDF and methimazole-treated ZDF (ZDF-methi) rats at 12 weeks of age. After collection, the blood samples were centrifuged (5 min, 14,000 r.p.m., 4℃) and serum samples were stored in liquid nitrogen until measurement. Serum T4 and corticosterone were measured using commercially available RIA kits from Monobind Incorporation (CA, USA) and IBL (Germany), respectively.
H&E staining and immunohistochemistry for Ki67 and DCX
For histological staining, ZLC, ZDF and ZDF-methi rats were perfused by a previous mentioned method (Hwang et al., 2008, 2009). In brief, adrenal glands were dehydrated with graded concentrations of alcohol for embedding in paraffin. Thereafter paraffin-embedded tissues were sectioned on a microtome (Leica, Wetzlar, Germany) into 3-µm coronal sections, and they were mounted into silane-coated slides. The sections were stained with hematoxylin and eosin (H&E) according to general protocol.
For immunohistochemistry, brains were cryoprotected by infiltration with 30% sucrose overnight. Thereafter, frozen tissues were serially sectioned on a cryostat (Leica) into 30 µm coronal sections and then the sections were collected into six-well plates containing PBS. Immunohistochemistry was performed under the same conditions in each group in order to examine whether the degree of immunohistochemical staining was accurate. Sections were sequentially treated with 0.3% hydrogen peroxide (H2O2) in PBS for 30 min and 10% normal goat or rabbit serum in 0.05 M PBS for 30 min. They were then incubated with diluted rabbit anti-Ki67 (1 : 1,000, Abcam, Cambridge, UK) or goat anti-DCX antibody (1 : 50, SantaCruz Biotechnology, Santa Cruz, CA, USA) overnight at room temperature and subsequently exposed to biotinylated goat anti-rabbit IgG or rabbit anti-goat IgG and streptavidin peroxidase complex (diluted 1 : 200, Vector, Burlingame, CA, USA). They were then visualized by staining with 3,3'-diaminobenzidine in 0.1 M Tris-HCl buffer (pH 7.2) and mounted on gelatin-coated slides. The sections were mounted in Canada Balsam (Kanto, Tokyo, Japan) following dehydration.
Data analysis
All measurements were performed in order to ensure objectivity in blind conditions, by two observers for each experiment, carrying out the measures of control and experimental samples under the same conditions.
For quantitative analysis of the number of Ki67 or DCX positive cells in the hippocampus, 15 section with 60 µm interval were selected from each animals according to anatomical landmarks corresponding to Bregma -3.00~-4.08 mm of rat brain atlas (Paxinos & Watson, 2007). The corresponding areas of the hippocampus were measured on the monitor at a magnification of 100×. Images of Ki67 or DCX-immunoreactive cells taken from dentate gyrus were obtained through a BX51 light microscope (Olympus, Tokyo, Japan) equipped with a digital camera (DP71, Olympus) connected to a PC monitor. The number of Ki67 or DCX positive cells in dentate gyrus was analyzed by Optimas 6.5 software (CyberMetrics, Scottsdale, AZ). In addition, dendritic complexity of DCX positive cells was analyzed using the accompanying software (NeuroExplore, MicroBrightField, Inc., VT,), calculating complexity including dendritic length and number of branches. Cell counts were obtained by averaging the counts from the sections taken from each animal: A ratio of the count was calibrated as %.
Statistical analysis
The GraphPad Prism (Ver 4.03) statistical analysis software was used for all data analysis. The data shown here represent the means of experiments performed for each experimental area. Differences among the means were statistically analyzed by one-way ANOVA test followed by Duncan's new multiple range method.
Results
Blood glucose, and serum levels of thyroid hormone and corticosterone
At 12 weeks of age, blood glucose levels were reported by our previous study (Hwang et al., 2009). Serum T4 levels in ZLC rats were 76.3 µg/dL. In ZDF rats, T4 levels were significantly increased to 10.31 µg/dL. In ZDF-methi rats, T4 levels were significantly decreased by 62% compared to that in the ZDF group. In this group, T4 levels were 3.95 µg/dL (Fig. 1A).
Serum corticosterone level in ZLC rats was 218.9 ng/mL. In ZDF rats, corticosterone level was significantly higher (461.7 ng/mL) than that in ZLC rats. In ZDF-methi rats, corticosterone level was significantly decreased (289.3 ng/mL) compared to that in ZDF rats (Fig. 1B).
Morphology of adrenal gland
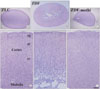
In ZLC, ZDF and ZDF-methi rats, morphologies of adrenal glands were differently found. First of all, the size of the adrenal medulla was markedly different among these groups. In ZDF rats, the adrenal medulla and the zona fasciculata of the adrenal cortex were significantly enlarged, but they were significantly decreased in ZDF-methi rats. Indeed, in the ZDF-methi rats, the size of adrenal gland was somewhat smaller than that in ZLC rats (Fig. 2A~F).
Changes in Ki67-immunoreactive cells in the dentate gyrus
In ZLC rats, some Ki67-immunoreactive nuclei were detected in the subgranular zone of the dentate gyrus (Fig. 3A). However, in ZDF rats, Ki67-immunoreactive nuclei were significantly decreased by 33.4% compared to those in ZLC rats (Fig. 3B and D). In ZDF-methi rats, Ki67-immunoreactive nuclei were significantly increased compared to those in ZDF rats (Fig. 3C). In this group, the number of Ki67-immunoreactive nuclei was slightly lower than that in ZLC rats (88.2% vs. ZLC rats) (Fig. 3D).
Changes in DCX-immunoreactive cells in the dentate gyrus
In ZLC rats, DCX-immunoreactive cells were detected in the subgranular zone of the dentate gyrus (Fig. 4A) and DCX had well-developed (tertiary) dendrites, which were extended into two-thirds of molecular layer of the dentate gyrus (Fig. 4B). In the ZDF rats, DCX-immunoreactive neuroblasts were significantly decreased in the dentate gyrus and fewer DCX-immunoreactive neuroblasts with tertiary dendrites were detected (Fig. 4C, D and G). In ZDF-methi rats, the number of DCX-immunoreactive neuroblasts with tertiary dendrites was increased compared to that in the ZDF rats and were similar to that in ZLC rats (Fig. 4E, F and G).
Discussion
The ZDF rat is a well-characterized genetic model of non-insulin-dependent (type 2) diabetes and obesity. ZDF rats have a defective leptin receptor (Zucker & Zucker, 1961) and diabetes typically appears between 7 and 10 weeks of age and is maintained for at least 6 months (Etgen & Oldham, 2000).
In this study, we observed that treatment with methimazole in ZDF rats decreased the serum blood glucose levels. In addition, methimazole treatment in ZDF rats significantly reduced the hypertrophy of adrenal medulla and zona fasciculata of adrenal cortex and serum corticosterone levels. These results are supported by previous studies that hypothyroidism in rats resulted in decreased adrenal weights and plasma concentrations of corticosterone (Tohei et al., 1991 and 1998; Tohei 2004).
We observed, in the present study, effects of methimazole on cell proliferation and neuronal differentiation in the subgranular zone of the hippocampal dentate gyrus in adult type 2 diabetic rats, because neurogenesis in the hippocampal dentate gyrus is associated with cognitive performance (Song et al., 2002; Siwak-Tapp et al., 2007; Aizawa et al., 2009). Ki67- and DCX-immunoreactive cells were markedly low in the dentate gyrus in ZDF rats, however, in methimazole treatment in ZDF rats, numbers of Ki67- and DCX-immunoreactive cells were similar to those ZLC rats.
Indeed, hippocampal neurogenesis is significantly decreased in diabetic animals (Jackson-Guilford et al., 2000; Beauquis et al., 2006; Hwang et al., 2008) and diabetic patients (Trudeau et al., 2004). Methimazole has dual actions on memory functions in healthy and diabetic rats. Hypothyroidism is also reported to impair cognition and memory in adult patients and animal models (Mennemeier et al., 1993; Wilcoxon et al., 2007) and to impair long-term potentiation in the rat hippocampus (Lee et al., 2003). In addition, thyroid dysfunction has been shown to influence acetylcholinesterase activity in both developing and adult rats (Carageorgiou et al., 2007). Furthermore, hypothyroidism reduces 5-bromo-deoxyuridine-positive cells (Ambrogini et al., 2005) and DCX-immunoreactive neuroblasts, which exhibit severely hypoplastic dendritic arborization (Montero-Pedrazuela et al., 2006). However, in ZDF rats, hypothyroidism alleviated the diabetic phenotypes and diabetes-induced reduction of DCX-immunoreactive cells. This effect may be associated with hypothyroidism-induced reduction of serum corticosterone levels. Treatment with antisense oligonucleotides directed against glucocorticoid receptor has been reported to restore normal fasting glucose levels in Zucker diabetic rats (Watts et al., 2005). In addition, lowering corticosterone prevented diabetes-induced impairment of learning and memory in insulin-deficient rats and insulin-resistant (db/db) mice (Stranahan et al., 2008). It has also been reported that corticosterone levels began to increase as a consequence of aging (Landfield et al., 1978), and lowering corticosterone from mid-age protected from the age-related decline in hippocampal neurogenesis and cognitive functions (Montaron et al., 2006). In addition, hypothyroidism induced by methimazole resulted in a significant decrease in the plasma concentrations of corticosterone (Weng et al., 2007), which has negative correlation with neurogenesis (Joëls 2007).
Exposure to elevated corticosterone reduces insulin receptor signaling in the brain (Piroli et al., 2007), and finally, the negative effect of diabetes on hippocampal plasticity may be attributable to an interaction between elevated glucocorticoids and insulin receptor signaling. In the present study, lowering corticosterone levels significantly increased Ki67- and DCX-immunoreactive cells in the dentate gyrus. This result is supported by a previous study that lowering corticosterone levels in diabetes could restore behavioral functions (Stranahan et al., 2008).
In conclusion, treatment with methimazole in ZDF rats, type 2 diabetic rats, significantly alleviated increases of serum corticosterone levels and enlargement of the adrenal gland. In addition, methimazole rescued the diabetes-induced reduction of Ki67- and DCX-immunoreactive cells. The reduction of diabetic phenotypes and rescue of cell proliferation and neuronal differentiation may be associated with hypothyroidism-related reduction of corticosterone.


 XML Download
XML Download