

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abstract
Background
Oxidative stress is thought to be one of the causative factors contributing to macrovascular complications in diabetes. However, the mechanisms of development and progression of diabetic vascular complications are poorly understood. We hypothesized that PKC-δ isozyme contributes to ROS generation and determined their roles in the critical intermediary signaling events in high glucose-induced proliferation of vascular smooth muscle (VSM) cells.
Methods
We treated primary cultured rat aortic smooth muscle cells for 72 hours with medium containing 5.5 mmol/L D-glucose (normal glucose), 30 mmol/L D-glucose (high glucose) or 5.5 mmol/L D-glucose plus 24.5 mmol/L mannitol (osmotic control). We then measured cell number, BrdU incorporation, cell cycle and superoxide production in VSM cells. Immunoblotting of PKC isozymes using phoshospecific antibodies was performed, and PKC activity was also measured.
Results
High glucose increased VSM cell number and BrdU incorporation and displayed significantly greater percentages of S and G2/M phases than compared to 5.5 mmol/L glucose and osmotic control. The nitroblue tetrazolium (NBT) staining in high glucose-treated VSM cell was more prominent compared with normal glucose-treated VSM cell, which was significantly inhibited by DPI (10 µM), but not by inhibitors for other oxidases. High glucose also markedly increased activity of PKC-δ isozyme. When VSM cells were treated with rottlerin, a specific inhibitor of PKC-δ or transfected with PKC-δ siRNA, NBT staining and NAD(P)H oxidase activity were significantly attenuated in the high glucose-treated VSM cells. Furthermore, inhibition of PKC-δ markedly decreased VSM cell number by high glucose.
Figures and Tables
 | Fig. 1Effect of high glucose on vascular smooth muscle cell proliferation (cell count) (A) and DNA synthesis (BrdU incorporation) of rat VSM cells (B).
Each VSM cells were incubated with media containing normal glucose (5.5 mmol/L D-glucose) or high glucose (30 mmol/L D-glucose) up to 72 h. At the end of incubation, cell number was measured by counting trypsinized cells with a hematocytometer and the incorporation of BrdU into VSM cells was examined using a microplate reader.
Data are obtained from three separate experiments and are expressed in the mean ± SE.
*p < 0.05 compared with normal glucose for 24 h.
†p < 0.01 compared with normal glucose for 72 h.
‡p < 0.01 compared with normal glucose for 72 h.
|
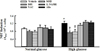 | Fig. 2Effect of various inhibitors such as DPI (10 µmol/L), NAC (10 µmol/L), SOD (500 U/mL), L-NAME (10 µmol/L), and allopurinol (10 µmol/L) on superoxide generation in normal and high glucose-treated VSM cells.
Data are shown as mean ± SE from 4 experiments in each group.
*p < 0.05 vs. normal glucose.
†p < 0.01 compared with corresponding values in each vehicle.
|
 | Fig. 3Effects of various inhibitors, such as DPI (10 µmol/L), SOD (500 U/mL), allopurinol (Allo, 100 µmol/L), rotenone (Rot, 100 µmol/L), L-NAME (NMA, 10 µmol/L), and indomethacin (IND, 10 µmol/L), NADH-stimulated superoxide production in normal and high glucose-treated VSM cells.
Data are shown as mean ± SE from 4 experiments in each group.
*p < 0.01 vs. normal glucose.
†p < 0.01 vs. vehicle.
|
 | Fig. 4Effects of high glucose on in vitro PKC activity in VSM cells.
Results were expressed as a mean percentage of 5.5 mmol/L D-glucose ± SE from 4 independent experiments.
*p < 0.01 vs. normal glucose.
|
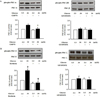 | Fig. 5Effects of high glucose on in vitro PKC activity and isozyme selective inhibition of PKC in VSM cells.
Confluent cultured VSM cells were treated with high glucose condition for 72 h. To verify the efficacy and selectivity of isozyme specific PKC inhibitors, VSM cells were treated with G6976 (1 µmol/L) and GF109203X (10 µmol/L) (A), rottlerin (3 µmol/L) and GF109203X (10 µmol/L) (B) for 1 h before incubation with high glucose for 72 h, lysed and immunoblotted by using PKC isozyme-specific antibodies recognizing phosphorylated forms of PKC-α and PKC-βII (A), PKC-δ and PKC-ζ (B). Total PKC isozymes in the immunoprecipitates were verified by immunoblotting with selective antibodies. The bar graph represents the mean ± SE of three separate experiments.
*p < 0.05 compared with high glucose without pretreatment with G6976.
†p < 0.01 compared with normal glucose.
‡p < 0.01 compared with high glucose without pretreatment with rottlerin.
|
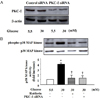 | Fig. 6Effect of selective PKC-δ inhibitor or PKC-δ siRNA on p38 MAP kinase. Quiescent VSM cells were pretreated with rottlerin (3 µmol/L) for 1 h or transfected with PKC-δ siRNA prior to stimulation with high glucose for 72 h. p38 MAP kinase activiy was assessed by immunoblotting with antibodies specific for phosphorylated p38 MAP kinase.
The bar graph represents the mean fold increases in three separate experiments.
Results are expressed as the mean ± SE.
*p < 0.01 compared with normal glucose.
†p < 0.01 compared with high glucose without treatment.
|
 | Fig. 7Effect of transient transfections of PKC-δ siRNA on cellular superoxide production in normal and high glucose-treated VSM cells. Superoxide production was measured by NBT reduction.
Results are presented as means ± SEM for three independent experiments.
*p < 0.05 vs. normal glucose.
†p < 0.01 vs. vehicle.
|
 | Fig. 8Effect of selective PKC inhibitor or PKC-δ siRNA on VSM cell proliferation (cell count). Quiescent VSM cellswere pretreated with rottlerin (3 µmol/L) for 1 h or transfected with PKC-δ siRNA prior to stimulation with high glucose for 72 h. At the end of incubation, cell number was measured by counting trypsinized cells with a hematocytometer.
Results are obtained from three separate experiments and are expressed in the mean ± SE.
*p < 0.01 compared with normal glucose.
†p < 0.05 compared with high glucose without treatment.
|
Table 1
Effect of High Glucose on the Cell Cycle Analysis Using Flow Cytometry

Cells were grown in 25-mm2 flasks and synchronized, and then the cell cycle was initiated by incubation with DMEM containing normal or high glucose for 72 h. The distribution of the cell cycle was examined as described in METHODS. Data are expressed as the percent distribution of cell cycle phases G0/G1, S, and G2/M in normal and high glucose. Similar results were obtained in three separate experiments.
*p < 0.01 compared with normal glucose.
![]()
References
1. DeFranzo RA, Ferrannini E. Insulin resistance. A multifaceted syndrome responsible for NIDDM, obesity, hypertension, dyslipidemia, and atherosclerotic cardiovascular disease. Diabetes Care. 1991. 14:173–194.
2. Ross R. Cell biology of atherosclerosis. Annu Rev Physiol. 1995. 57:791–804.
3. Tanner FC, Boehm M, Akyurek LM, San H, Yang ZY, Tashiro J, Nabel GJ, Nabel EG. Differential effects of the cyclin-dependent kinase inhibitors p27(KIP1), p21(CIP1), and 16(Ink4) on vascular smooth muscle cell proliferation. Circulation. 2000. 10:2022–2025.
4. Bruemmer D, Berger JP, Liu J, Kintscher U, Wakino S, Fleck E, Moller DE, Law RE. A non-thiazolidinedione partial peroxisome proliferator-activated receptor ligand inhibits vascular smooth muscle cell growth. Eur J Pharm. 2003. 466:225–234.
5. Brownlee M. Biochemistry and molecular cell biology of diabetic complications. Nature. 2001. 414:813–820.
6. Griendling KK, Taubman MB, Akers M, Mendlowitz M, Alexander RW. Characterization of phosphatidylinositol-specific phospholipase C from cultured vascular smooth muscle cells. J Biol Chem. 1991. 266:15498–15504.
8. Beckman JA, Creager MA, Libby P. Diabetes and atherosclerosis: epidemiology, pathophysiology, and management. JAMA. 2002. 87:2570–2581.
9. Creager MA, Luscher TF, Cosentino F, Beckman JA. Diabetes and vascular disease: pathophysiology, clinical consequences, and medical therapy. Part I. Circulation. 2003. 108:1527–1532.
10. Barsness GW, Peterson ED, Ohman EM, Nelson CL, DeLong ER, Reves JG, Smith PK, Anderson RD, Jones RH, Mark DB, Califf RM. Relationship between diabetes mellitus and long-term survival after coronary bypass and angioplasty. Circulation. 1997. 96:2551–2556.
11. King GL, Kunisaki M, Nishi N, Inoguchi T, Shiba T, Xia P. Biochemical and molecular mechanisms in the development of diabetic vascular complications. Diabetes. 1996. 45:Suppl 3. S105–S108.
12. Lee HS, Son SM, Kim YK, Hong KW, Kim CD. NAD(P)H oxidase participates in the signaling events in high glucose-induced proliferation of vascular smooth muscle cells. Life Sci. 2003. 72:2719–2730.
13. Inoguchi T, Li P, Umeda F, Yu HY, Kakimoto M, Imamura M, Aoki T, Etoh T, Hashimoto T, Naruse M, Sano H, Utsumi H, Nawata H. High glucose level and free fatty acid stimulate reactive oxygen species production through protein kinase C-dependent activation of NAD(P)H oxidase in cultured vascular cells. Diabetes. 2000. 49:1939–1944.
14. Graier WF, Grubenthal I, Dittrich P, Wascher TC, Kostner GM. Intracellular mechanism of high D-glucose-induced modulation of vascular cell proliferation. Eur J Pharmacol. 1995. 294:221–229.
15. Nakamura J, Kasuya Y, Hamada Y, Nakashima E, Naruse K, Yasuda Y, Kato K, Hotta N. Glucose-induced hyperproliferation of cultured rat aortic smooth muscle cells through polyol pathway hyperactivity. Diabetologia. 2001. 44:480–487.
16. Natarajan R, Gonzales N, Xu L, Nadler JL. Vascular smooth muscle cells exhibit increased growth in response to elevated glucose. Biochem Biophys Res Commun. 1992. 187:552–560.
17. Sheetz MJ, King GL. Molecular understanding of hyperglycemia's adverse effects for diabetes complications. JAMA. 2002. 288:2579–2588.
18. Igarashi M, Wakasaki H, Takahara N, Ishii H, Jiang ZY, Yamauchi T, Kuboki K, Meier M, Rhodes CJ, King GL. Glucose or diabetes activates p38 mitogen-activated protein kinase via different pathways. J Clin Invest. 1999. 103:185–195.
19. Haller H, Baur E, Quass P, Behrend M, Lindschau C, Distler A, Luft FC. High glucose concentrations and protein kinase C isoforms in vascular smooth muscle cells. Kidney Int. 1995. 47:1057–1067.
20. Konishi H, Tanaka M, Takemura Y, Matsuzaki H, Ono Y, Kikkawa U, Nishizuka Y. Activation of protein kinase C by tyrosine phosphorylation in response to H2O2. Proc Natl Acad Sci USA. 1997. 94:11233–11237.
21. Lagaud GJ, Masih-Khan E, Kai S, van Breemen C, Dube GP. Influence of type II diabetes on arterial tone and endothelial function in murine mesenteric resistance arteries. J Vasc Res. 2001. 38:578–589.
22. Park JY, Takahara N, Gabriele A, Chou E, Naruse K, Suzuma K, Yamauchi T, Ha SW, Meier M, Rhodes CJ, King GL. Induction of endothelin-1 expression by glucose: an effect of protein kinase C activation. Diabetes. 2000. 49:1239–1248.
23. Suzuma K, Takahara N, Suzuma I, Isshiki K, Ueki K, Leitges M, Aiello LP, King GL. Characterization of protein kinase C isoform's action on retinoblastoma protein phosphorylation, vascular endothelial growth factor-induced endothelial cell proliferation, and retinal neovascularization. Proc Natl Acad Sci USA. 2002. 99:721–726.
24. Fukumoto S, Nishizawa Y, Hosoi M, Koyama H, Yamakawa K, Ohno S, Morii H. Protein kinase C inhibits the proliferation of vascular smooth muscle cells by suppressing G1 cyclin expression. J Biol Chem. 1997. 272:13816–13822.
25. Ueda Y, Hirai S, Osada S, Suzuki A, Mizuno K, Ohno S. Protein kinase C activates the MEK-ERK pathway in a manner independent of Ras and dependent on Raf. J Biol Chem. 1996. 271:23512–23519.
26. Guo JH, Wang H, Malbon CC. Conditional, tissue-specific expression of Q205L Gi2 in vivo mimics insulin activation of c-Jun N-terminal kinase and p38 kinase. J Biol Chem. 1998. 273:16487–16493.
27. Sweeney G, Somwar R, Ramlal T, Volchuk A, Ueyama A, Klip A. An inhibitor of p38 mitogen-activated protein kinase prevents insulin-stimulated glucose transport but not glucose transporter translocation in 3T3L1 adipocytes and L6 myotubes. J Biol Chem. 1999. 274:10071–10078.
28. Igarashi M, Yamaguchi H, Hirata A, Tsuchiya H, Susa S, Tominaga M, Daimon M, Kato T. Insulin activates p38 mitogen-activated protein (MAP) kinase via a MAP kinase kinase (MKK) 3/MKK 6 pathway in vascular smooth muscle cells. Eur J Clin Invest. 2000. 30:668–677.
29. MacFarlene WM, Smith SB, James RFL, Clifton AD, Doza YN, Cohen P, Docherty K. The p38/reactivating kinase mitogen-activated protein kinase cascade mediates the activation of the transcription factor insulin upstream factor 1 and insulin gene transcription by high glucose in pancreatic β-cells. J Biol Chem. 1997. 272:20936–20944.
30. Stein B, Brady H, Yang MX, Young DB, Barbosa MS. Cloning and characterization of MEK6, a novel member of the mitogen-activated protein kinase cascade. J Biol Chem. 1996. 271:11427–11433.
31. Krump E, Sanghera JS, Pelech SL, Furuya W, Grinstein S. Chemotactic peptide N-formyl-met-leu-phe activation of p38 mitogen-activated protein kinase (MAPK) and MAPK-activated protein kinase-2 in human neutrophils. J Biol Chem. 1997. 272:937–944.
 XML Download
XML Download