

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Asthma is an inflammatory disease of airways, which is characterized by hyperreactivity of trachea and bronchi in response to a group of stimulators [1]. Asthmatics are suffering from narrowing airways with clinical manifestations like cough, wheeze and respiratory distress. Asthma mostly occurs in childhood. Genetic factors, viruses, allergens and job are important players in development of asthma [2, 3, 4].
IgE mediated immune responses are the main pathologic reactions in asthma. Significant inflammation exists in bronchial biopsy samples of asthmatic patients even in mild cases and these inflammatory changes affect entire airways [5, 6].
In allergic asthma, the possibility of a familial history of rhinitis, urticaria and eczema, and elevated levels of IgE antibody exists. However, many asthmatics have not any familial history of allergy and their serum IgE levels are normal. Therefore, this kind of asthma is named nonallergic asthma [2, 7, 8]. Moreover, based on the pattern of infiltrating inflammatory cells, asthma has been divided into two groups: eosinophilic asthma and noneosinophilic asthma [9].
In asthmatics, CD4+ T cells are increased in allergic airways with dominancy of T helper (Th) 2 cell population [10]. However, it seems that the role of cytokines produced by Th25, Th22, Th17, and Th9 cells might be important in immunopathogenesis of asthma.
A new subtype of CD4+ T cells which is called Th9, secrets high levels of interleukin-9 (IL-9). This cytokine can change the expression pattern of other inflammatory cytokines in lung epithelial cells [11, 12]. IL-9 stimulates the proliferation of activated T cells, promotes the proliferation and differentiation of mast cells (MCs) and increases the production of IgE by B cells [13, 14, 15].
Th17 cells, by producing IL-17, have critical roles in inflammatory and autoimmune diseases such as rheumatoid arthritis, multiple sclerosis and periodontitis [16]. In addition, Th17 cells may have a role in severe asthma and chronic obstructive pulmonary disease [15, 17, 18].
IL-22, a member of IL-10 super family, initiates innate immune responses against bacterial pathogens especially in epithelial cells such as respiratory and gut epithelial cells [19, 20]. IL-22, which is the signature cytokine of newly identified Th22 cells, exerts its role as an inflammatory mediator and a stimulator of acute phase proteins production in inflammatory conditions such as psoriasis, rheumatoid arthritis and Crohn's disease [18, 21, 22]. Several researches have suggested that IL-22 has a key role in the pathogenesis of allergic inflammation of the airways [23].
Another recently identified subtype of CD4+ helper T cells is Th25 as it is the main lymphoid source of IL-25 production. IL-25 has the potential of deviating immune response towards Th2 pattern in allergic asthma and recent studies have clarified its role in mediating allergic inflammation and infiltration of eosinophils and macrophages in airways of asthmatics patients [24, 25, 26].
A new subset of lymphocytes which is called innate lymphoid cell (ILC) represents a family of cell types which do not express lymphoid lineage markers (Lin-), but are CD161 and the IL-7 receptor-α (CD127) positive [27]. The importance of these cells is in their ability to produce significant amount of cytokines similar to different subtypes of CD4+ T cells. These cells are classified as ILC1, ILC2, and ILC3. Among ILC subtypes, one of them, ILC2, produces type 2 cytokines such as IL-4, IL-9, and IL-13 [28].
The aim of this study was to evaluate the expression of newly identified helper T cells signature cytokines that were likely to have important roles in the pathogenesis of asthma. Comparing their expression in sputum and serum samples in addition to counting ILC cells as another important source of such inflammatory cytokines were the other missions of the current study.
Go to : 
MATERIALS AND METHODS
Subjects
Twenty-three patients with asthma who had been referred to allergy and asthma clinic of Alzahra Hospital, Isfahan, Iran, were diagnosed and selected according to the Global Initiative for Asthma criteria. Patients with severe asthma were defined as previously described [29]. Informed consent was obtained from all studied subjects. Study was approved by the local ethical committee of the Isfahan University of Medical Sciences, Iran. None of the patients had taken the medication at least one week before sampling. The controls were 23 healthy individuals matched for age and sex and with no history of allergy, asthma and other inflammatory diseases at the time of sampling. The Characteristics of the asthmatic patients and control group are summarized in Table 1. From each individual, 10-mL peripheral blood was collected, half with ethylenediaminetetraacetic acid (EDTA) and half without any anticoagulant. Subjects were stimulated with NaCl concentration gradient using nebulizer (3%, 4%, and 5%) and the sputum samples were collected in the sterile tubes.
RNA extraction and cDNA synthesis
RNA was extracted using the Thermo Scientific Gene JET RNA Purification Kit (Thermo Fisher Scientific Inc., Waltham, MA, USA). Sputum RNA was extracted using RNX-PLUS reagent (CinnaGen, Karaj, Iran). RNA gel electrophoresis was performed to assess RNA quality, and samples were included in the study if 28S ribosomal RNA bands were present with intensity that is approximately twice that of the 18S RNA band. Finally, RNA concentration was measured by Nanodrop (Biochrom WPA Biowave II, Biochrom, Cambridge, UK) spectrophotometer. Total RNA was converted into the cDNA using Aid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific Inc.) as instructed by the manufacturer.
Real-time quantitative transcription-polymerase chain reaction
The resulting transcripts were then quantified by the real-time quantitative polymerase chain reaction (PCR) on a StepOneplus real-time DNA amplification system (Applied Biosystems, Foster City, CA, USA) using SYBR Premix Ex Taq II kit (Takara Bio Inc., Otsu, Japan) according to the manufacturer's instructions. Predesigned primers (QuantiTact primer Assay; Qiagen, Venlo, The Netherlands) specific for amplification of IL-9, IL-17, IL-22, and IL-25 were applied. For each sample, transcript quantity was normalized to the amount of beta-actin (ACTB) expression. Amplification was carried out in a total volume of 20 µL for 40 cycles of 30 seconds at 95℃ and 30 seconds at 60℃.
Cell staining and flow cytometric analysis
Collected blood samples with EDTA were lysed using a RBC lysis solution (CMG, Isfahan, Iran) according to the manufacturer's instructions. Staining for flow cytometry was performed using the fluorescein isothiocyanate-conjugated anti-CD3, anti-CD19 (eBiosciences, San Diego, CA, USA) and anti-CD56 antibody (NCAM) (Santa Cruz Biotechnology Inc., Dallas, TX, USA), all for lineage exclusion, as well as with peridinin chlorophyll protein (PerCp)-conjugated antihuman CD127 (IL-7Rα) antibody (R&D systems Inc., Minneapolis, MN, USA) and phycoerythrin-conjugated Anti-Human CD161 antibody (eBiosciences) for detecting ILCs. A minimum of 100,000 lymphocytes were acquired per sample. Prepared samples were acquired on a FACScan and data were analyzed with the CellQuest-Pro software (BD Biosciences, San Jose, CA, USA).
Immunoblotting
In order to identify the allergens to which the patients were sensitive,we used 300 µL of patient serum samples was checked with Alleisa screen kit (Mediwiss Analytic GmbH, Moers, Germany) having 30 common allergens put on a nitrocellulose paper band.
Statistical analysis
The results were statistically analyzed with the independent sample t test using SPSS ver. 16.0 (SPSS Inc., Chicago, IL, USA). Results are expressed as mean ± standard deviation, and p values <0.05 are considered as significant.
Go to :
RESULTS
Cytokines were increased in the serum and sputum samples of asthmatics
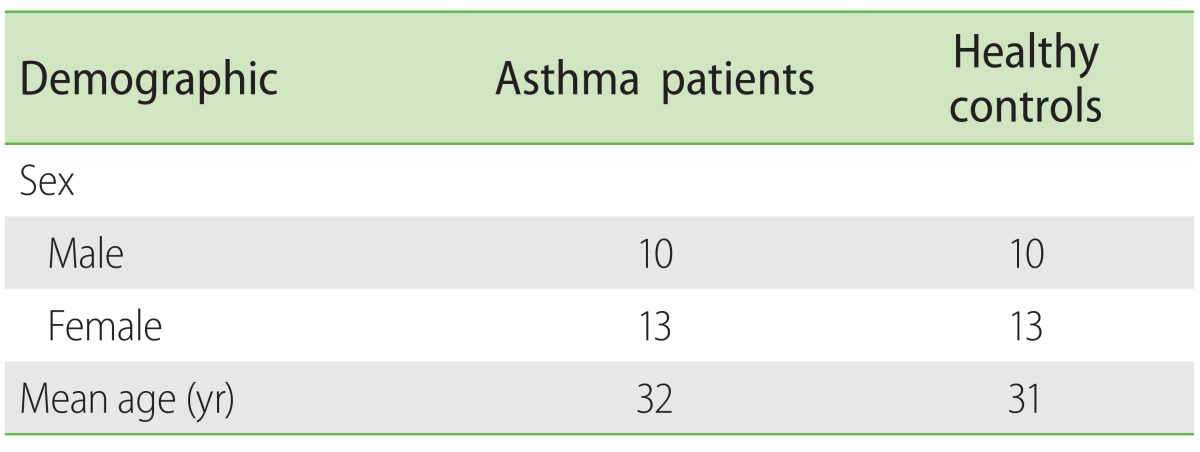
Experiments showed that gene expression of IL-9, IL-17, IL-22, and IL-25 cytokines in serum samples of the patients with asthma were hundred folds more than normal controls (Fig. 1A) and this increase was quite significant (p < 0.05). Moreover, a significant increase (p < 0.05) in the expression levels of the same cytokines was observed in the sputum samples of the patients in comparison with normal individuals (Fig. 1B). It also revealed that the expression levels of the cytokines in the serum samples of the asthmatics were significantly more than their levels in patient's sputum samples (p < 0.05) (Fig. 1C). However, no significant difference between the cytokines expression levels in serum samples and sputum samples of the controls was seen except in expression level of IL-22, which was significant (Fig. 1D).
 | Fig. 1Average expression of interleukin (IL) 9, IL-17, IL-22, and IL-25 cytokines in asthma patients and normal controls, using quantitative transcription-polymerase chain reaction. (A) Average expression of IL-9, IL-17, IL-22, and IL-25 cytokines in serum samples of cases was significantly increased compared with controls (p < 0.05). (B) Average expression of IL-9, IL-17, IL-22, and IL-25 cytokines in sputum samples of cases was significantly higher than controls (p < 0.05). (C) Comparison of the expression of IL-9, IL-17, IL-22 and IL-25 cytokines in serum and sputum samples of the asthmatics showed that the increase is much greater in serum samples (p < 0.05). (D) Comparison of the expression of IL-9, IL-17, and IL-25 cytokines in serum and sputum samples of the normal controls revealed no significant differences. However it was significant for IL-22 expression level. *p < 0.05, statistical significance.
|
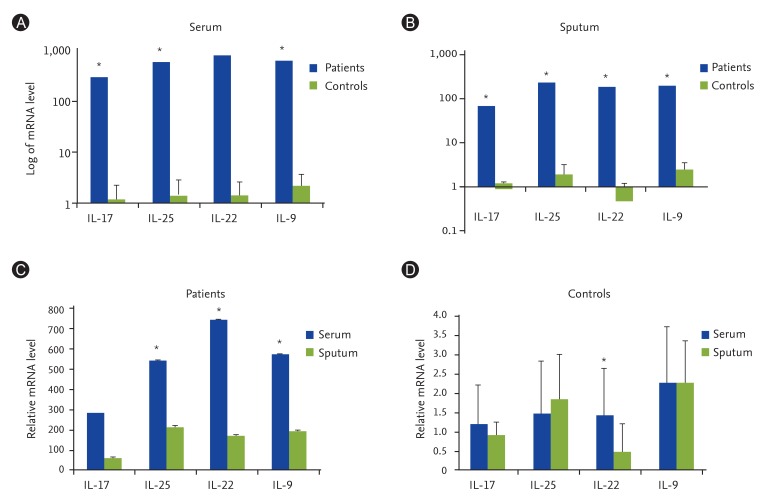
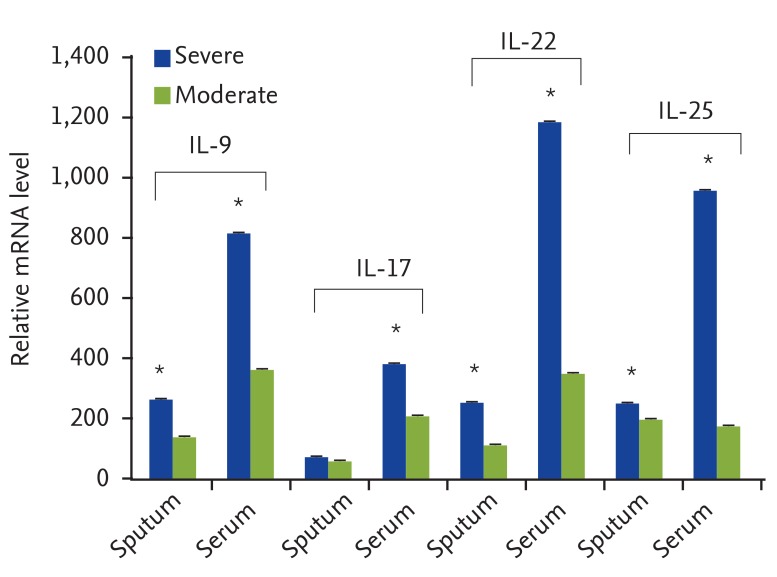
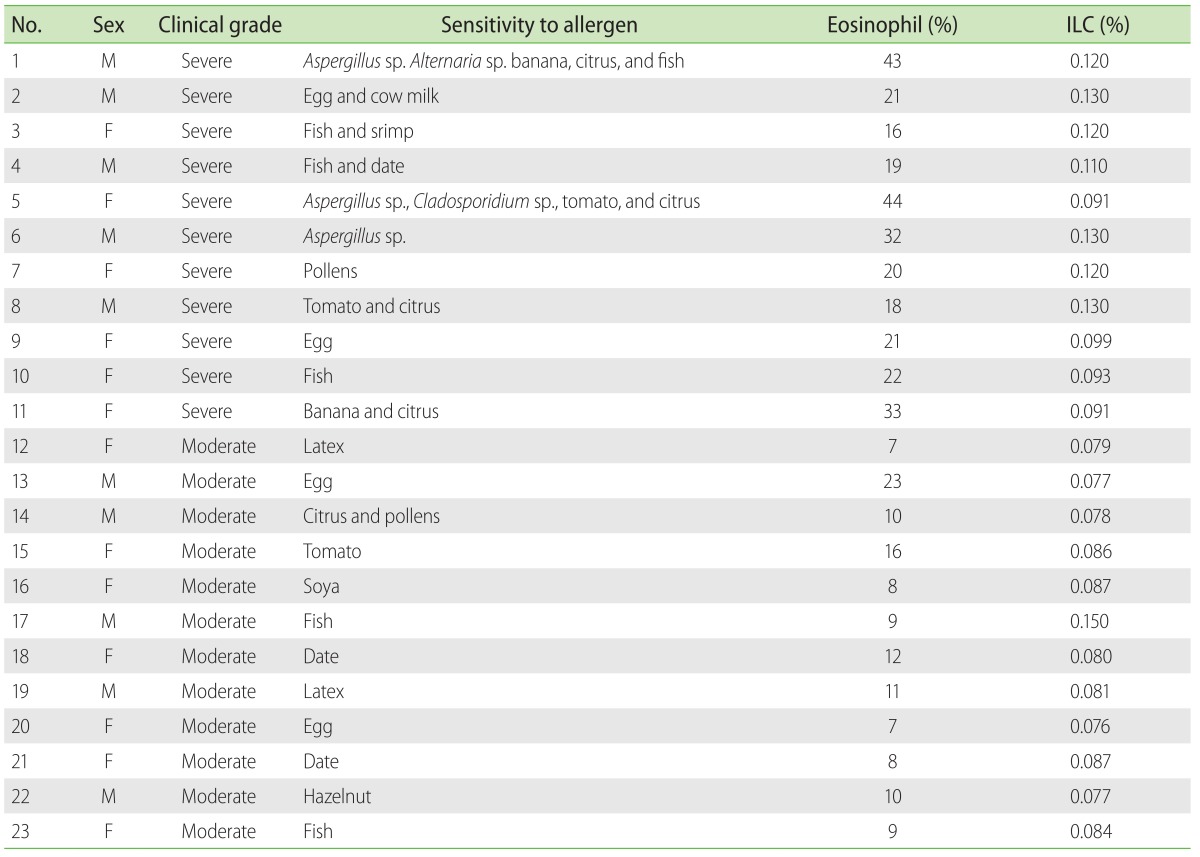
Table 2 shows the immunoblotting results as well as eosinophil and ILC percentages of the asthmatic cases. Of the total 23 asthma patients, 11 were suffering from severe asthma. Immunoblotting results showed that the most of the patients with severe asthma were sensitive to more than one allergen. Furthermore, average percentages of eosinophils in the peripheral blood of severe and moderate forms of asthma were 26% ± 10.06% and 11% ± 4.57%, respectively (Table 2). Accordingly, it is shown that the transcript levels of IL-9, IL-17, IL-22, and IL-25 cytokines in serum samples of severe asthma, in comparison with moderate asthma, were significantly augmented (p < 0.05). Although the levels of IL-9, IL-22, and IL-25 were significantly higher in sputum samples of severe asthma, this difference was not statistically significant for IL-17 expression (Fig. 2).
 | Fig. 2Comparing the average cytokines expression in serum and sputum samples of patients with severe and moderate asthma. Transcript levels of IL-9, IL-17, IL-22, and IL-25 cytokines in serum samples of severe asthma is significantly higher in comparison with moderate asthma (p < 0.05). In sputum samples, only IL-17 expression is not significantly more in severe asthma. *p < 0.05, statistical significance.
|
ILC count was increased in the peripheral blood of asthma patients
In order to further investigate the relationship between ILCs and asthma patients, we counted ILCs, represented by Lin-/CD127+/CD161+ phenotype, in the peripheral blood of the cases and controls using flow cytometry. As ILCs are morphologically similar to lymphocytes, in the histograms of peripheral blood cell analysis, lymphoid cells were gated (R1) (Fig. 3A). Then, in order to exclude the T cells, B cells, and natural killer cells, which express CD3, CD19, and CD56, respectively, we analyzed the lymphoid cells (gated as R1) for the absence of these markers (e.g., Lin-) (Fig. 3B). After gating on Lin- cells, a population of CD127+/CD161+ cells distinct from the conventional lymphoid cells was present (Fig. 3C, D). Thus, the Lin-/CD127+/CD161+ cells most likely represented for ILCs population. These results showed that Lin-/CD127+/CD161+ ILCs were significantly higher in the peripheral blood of the asthma patients compared to the healthy individuals (0.099% ± 0.024% vs. 0.036% ± 0.0215, respectively; p < 0.05). In addition, average percentage of ILCs in the peripheral blood of patients with severe and moderate forms of asthma was 0.11% ± 0.028% and 0.08% ± 0.02%, respectively (p < 0.05). However, there was not significant correlation between ILC% and studied cytokine levels.
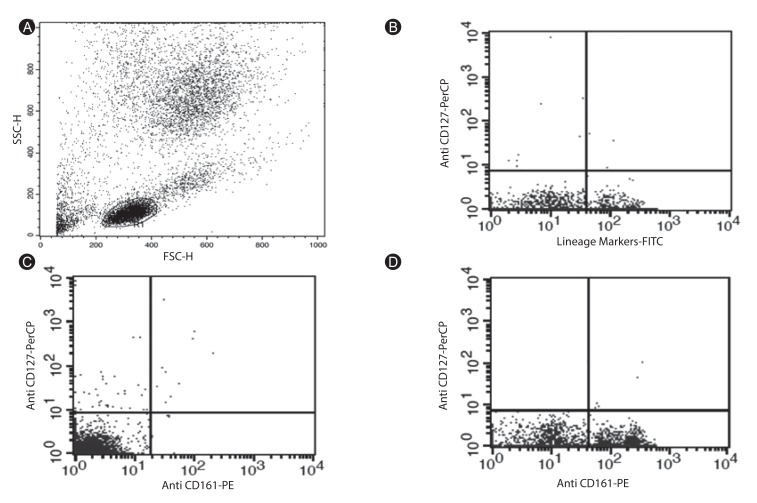 | Fig. 3Flow cytometry analysis of the Lin-CD127+CD161+ innate lymphoid cells (ILCs) in the peripheral blood of the asthma patients compared with the healthy individuals. (A) Forward scatter/side scatter cannels diagram of peripheral blood cells shows gated area for lymphoid cells. (B) Flow cytometry analysis of the peripheral blood samples in which T cells, B cells, and natural killer cells are excluded as one population using the fluorescein isothiocyanate-conjugated anti-CD3, anti-CD19, and anti-CD56 antibodies. Lin-/CD127+ are seen in upper left area. (C) The diagram shows the Lin-CD127+CD161+ ILCs count in an asthma patient, which appears in upper right area. (D) Diagram of the Lin-CD127+CD161+ ILCs count in a normal control. These cells are in upper right area.
|
Go to :
DISCUSSION
Asthma is defined as a clinical syndrome of respiratory system that initiates by viral infections of the upper respiratory system, environmental allergens or other irritants. Its main features are nonspecific bronchial hypersensitivity and inflammation of the respiratory tract [30]. Correlation of more than one hundred genes with asthma has been shown so far, but most of these studies need to be replicated and confirmed on wider scale and in different populations [31].
Although, IL-9 is now known as a potent and multifunctional factor, its exact role in inflammation and autoimmune diseases is still unclear. Therefore, several studies have been done on IL-9 and its correlation with respiratory diseases. Current study showed that the transcript levels of IL-9 in the blood and sputum samples of asthmatic patients were increased significantly in comparison with healthy subjects (p < 0.05).
Independent studies have shown that the elevation of IL-9 transcript levels in manipulated mice results in important changes in their lungs, such as lymphocytic inflammation, mucus hypersecretion, increased frequency of MCs in the lungs, eosinophils and neutrophils recruitment to the airways, and destruction of the subepithelial collagen; all these are representing the important role of IL-9 in chronic allergic inflammation [32, 33, 34]. One study has shown that the transcript levels of IL-9 significantly have increased in the tissue samples of asthmatic airways, not only in comparison with normal samples, but also in comparison with chronic bronchitis. On the other hand, several studies have shown that IL-9 is a potent activator of MCs, B cells and neutrophils and stimulates the secretion of mucus in the lungs [35, 36]. Our results are consistent with previous studies performed in other populations and in fact, this study confirms the results of previous studies on IL-9.
Th17 lymphocytes have been shown as a T-cell subset that produces high levels of IL-17 in response to inflammatory cytokines such as IL-1β, IL-6, and IL-23 [37, 38, 39]. Our results showed that the transcript levels of IL-17 significantly increased in the peripheral blood and sputum of asthmatic patients compared with control subjects.
One study in 2001 had shown that the expression of IL-17 significantly was increased in the sputum and bronchoalveolar lavage of asthmatic patients in comparison with controls [40]. Bullens et al. [39] indicated that IL-17 mRNA was significantly increased in sputum of asthmatic patients compared with healthy controls. Also, a recent study in 2013 has reported that the level of IL-17 gene expression is significantly increased in the blood of children with severe asthma compared with mild and moderate forms of asthma [41]. Our findings are consistent with the results of these studies.
We also found that there are significant differences between IL-17 expressions in severe asthma compared with moderate asthma. Bullens et al. [39] have shown that IL-17 gene expression is markedly more in severe asthmatic patients than in mild allergic form. Although, they didn't compare severe with moderate forms of asthma, their report is still consistent with our result. These results suggest that the elevation of IL-17 gene expression can play important roles in the incidence of asthma, presumably in a way similar to its role in other inflammatory diseases. Thus, it could be considered as a potential therapeutic target for asthma.
IL-22 is produced by Th17, ILCs and particularly by Th22 cells [42]. Inflammatory cytokines like IL-6 and tumor necrosis factor-α make Th22 cells to produce IL-22. Depending on the environmental factors, IL-22 can exert proinflammatory or anti-inflammatory functions [43]. We indicated that the transcript level of IL-22 is significantly increased in the peripheral blood and sputum samples of asthmatic patients compared with control subjects.
Two independent studies have demonstrated that the transcript level of IL-22 is increased in blood samples of asthmatic patients compared with healthy subjects [44, 45]. Zhao et al. [46] have reported that serum level of IL-22 is increased in patients with severe asthma compared to moderate asthma. Our results confirm these findings. According to our survey in literature, no report was found for IL-22 expression level in sputum samples of asthmatic patients in order to compare with our results.
A number of studies have shown anti-inflammatory effects of IL-22 in inflammatory airways diseases. Schnyder et al. [47] have reported that IL-22 may prevent inflammatory responses by modulation of dendritic cell functions in asthmatic mouse models. In addition, Takahashi et al. [43] have demonstrated that applying anti-IL-22 antibody resulted in increased production of inflammatory cytokines and eosinophil infiltration in a murine model of asthma. According to these and our findings on IL-22 levels in asthmatic patients, one can suppose that the elevation of IL-22 might be a reaction of the body in order to resist against the elevation of other inflammatory cytokines to dampen their destructive effects. However, its exact roles and mechanisms need to be confirmed by specific investigations.
IL-25 (IL-17E) is a cytokine secreted by epithelial cells, MCs, ILCs and especially by Th25 cells and induces the production of IL-4, IL-5, IL-9, and IL-13 cytokines in multiple tissues [15, 48]. The Present study showed that, in compare with controls, the transcript level of IL-25 is significantly higher in the peripheral blood and sputum samples of asthmatic patients.
Gregory et al. [49] have shown that when mice are exposed to allergens, the transcript level of IL-25 increases in the lung epithelial cells and these mice shows the manifestation of allergic asthma. They also have reported that injection of IL-25 to mice results in elevation of the transcript levels of IL-4, IL-5, IL-13 as well as serum IgE level, eosinophilia and pathological changes in airways. It's demonstrated that, by inducing histological changes in lung, IL-25 production by lung epithelial cells is required for the development of the disease in airways [50]. In one study, it is shown that neutralization of IL-25 leads to reduction of inflammation in airways and decreasing of type 2 cytokines in mouse models [26]. Our results are consistent with the results of these studies and confirm that the elevation of IL-25 may play an important role in asthma. This effect is probably similar to what IL-25 exerts in other inflammatory diseases.
In the present study, the gene expression levels of the cytokines have been measured both in peripheral blood and in sputum samples. We showed that the mRNA levels of IL-9, IL-17, IL-22, and IL-25 cytokines are much more in the peripheral blood than in the sputum samples of the patients. Thus, measurement of these cytokines in the peripheral blood has higher diagnostic value in comparison with sputum sample of asthmatic patients. This difference may be due to the greater amount of the cells that produce these cytokines in the peripheral blood than in their sputum, more degradation of mRNA in RNase-rich environment of sputum and also inappropriate living conditions for inflammatory cells in mucosal secretions of sputum compared with the blood. We did not find similar reports regarding to this finding to compare with our findings.
ILCs, as a new family of immune cells, produce different cytokines such as IL-5, IL-13, and IL-9 [27, 28]. We showed that the ILCs count in the asthmatic patients is significantly higher than the controls. Average percentage of these cells was 0.036% in the healthy individuals and this is consistent with the study of Mjosberg et al. [27] who had reported this frequency as 0.01-0.03% in the normal population. Regarding to the ILC percentage in asthmatics, we didn't find similar reports to compare with our findings. Both elevations in ILCs and eosinophils counts and augmentation in inflammatory cytokines in asthmatic patients could show that there is cooperation among ILCs, eosinophils and helper T lymphocytes in ignition and maintenance of allergic inflammation.
In conclusion, the results presented in this study show that IL-9, IL-17, IL-22, and IL-25 are potentially related to asthma and severity of asthmatic symptoms. These cytokines and/or their receptors could be considered as potential therapeutic targets for controlling the disease process. Also, enhancement of ILCs as another important source of inflammatory cytokines could help us to have a better understanding of immunopathology of asthma and to designing of new therapeutic strategies.
Go to :
 XML Download
XML Download