

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Theophylline is a methylxanthine derivative with a narrow therapeutic index, and at a serum concentration between 5-15 µg/mL, offers an effective and safe means of controlling the symptoms of asthma and bronchospasms caused by diseases such as COPD [1]. Moreover, there is a close relationship between the serum concentration of theophylline and its toxic and therapeutic effects [2]. Theophylline is metabolized primarily in the liver by one or more cytochrome P450 enzymes in humans, whereby theophylline is N-demethylated to 3-methylxanthine (3MX) and 1-methylxanthine (1MX), the latter of which is oxidized rapidly by xanthine oxidase to 1-methyluric acid [3]. Theophylline is also converted into 1,3-dimethyluric acid (DMU) via 8-hydroxylation which accounts for about half of its clearance when orally administered [4]. CYP1A2 is the major enzyme responsible for the N-demethylation and hydroxylation of theophylline, although CYP2E1 also plays a minor role in the metabolism of theophylline by catalyzing its 8-hydroxylation [4-6]. There are factors which is likely to affect theophylline metabolism by these enzymes, such as antiepileptics, oral contraceptives, smoking habit and caffeine, so on.
CYP1A2 and CYP2E1 are expressed in human liver, although large individual differences have been reported in hepatic expressions of CYP1A2 and CYP2E1 [7-11]. Moreover, several studies have suggested a relationship between the rate of drug metabolism and the genotype of the CYP1A2 gene. For example, large interindividual differences in CYP1A2 and CYP2E1 gene expressions, and in their capacities to metabolize xenobiotics, including theophylline [7, 8, 12-14], and large interethnic variations have been described [15, 16].
In an effort to understand the mechanisms underlying the regulation and induction of CYP1A2, its 5'-flanking region was examined [17, 18]. Recently Obase et al. [19] reported that theophylline metabolism levels were lower in patients with the A allele at -3860G>A, particularly in young asthmatics. Similar studies have been performed by Nakajima et al. [20] and Welfare et al. [21] The Welfare group found no significant differences in the DNA sequences of rapid and late caffeine metabolizers in the region -2,521 base pairs (bp) upstream from exon 7 of the CYP1A2 gene, whereas, in contrast, the Nakajima group found that a polymorphism at the 5'-flanking region of the CYP1A2 gene is related to caffeine metabolism in current smokers.
Therefore, the purpose of this study is to examine the relationships between theophylline metabolism and genetic variants of the theophylline metabolizing enzymes (CYP1A2 and CYP2E1) in Korean non-smoking asthmatic patients. In addition, we investigated whether any transcription factor related to modulation of the activities of these enzymes.
MATERIALS AND METHODS
Study subjects
A total of 210 patients with bronchial asthma, aged 34 to 79 years, who regularly visited Seoul National University Hospital, Seoul, Korea from January to September in 2002 and who had been taking a sustained-release formulation of theophylline (Etheophyl; Lindopharm GmbH, Germany: Theoclear; H-Pharm, Korea) were included. Most of them co-administrated drugs with theophylline, such as inhaled corticosteroids (96.5%), leukotrienes (1.2%), and systemic steroids (16.3%).
121 Patients were excluded according to the following criteria: 27 patients whose accompanied disease were excluded, 1 patient with hepatic or renal dysfunction, 1 patient with hypo/hyperthyroidism, 2 patients with congestive heart failure, 21 poor compliances, 9 patients with inadequately classified atopy with an indeterminable asthma type, 8 patients with no historical data of smoking habit and 4 patients whose blood sample was unable to be analyzed. In addition, the following patients were excluded, those morbidly obese, those on medication likely to affect theophylline metabolism (e.g.antiepileptics), current smokers, habitual users of caffeine (coffee, tea and soft drinks), and those who regularly took oral contraceptives. Finally, 89 asthma patients were included (Table 1). The study protocol was approved by ethics committees of Seoul National University Hospital, and all participants gave informed consent.
Genotyping
We screened 5'-flanking region of CYP1A2 and CYP2E1 including transcription factor binding site and polymorphisms which is known to affect the genetic regulation and function [18, 22]. To determine genotypes, genomic DNA was extracted from 10 mL of whole blood in ACD tubes using a Wizard Genomic DNA Purification Kit (Promega Madison, USA). DNA samples were dissolved in 10 nM Tris/1 nM EDTA (pH 8.0) and stored at -20℃ before polymerase chain reaction (PCR) amplification. Four regions of CYP1A2 including the 5'-flanking region and intron 1, and four regions of CYP2E1 including the 5'-flanking region were amplified by PCR. Primers for these eight sites were designed according to published sequences (GenBank Accession number AF 253322; CYP1A2, Gene card Accession number NM_000773; CYP2E1). Oligo Primer Analysis Software Version 5 (Molecular Biology Insights Inc, USA) was used to design the PCR primer pairs described in Table 2. PCR fragments were analyzed in 2% agarose/1× TAE gels. PCR mixtures (50 µL) were comprised of 1.25 U of Taq Polymerase (Boehringer Mannheim, Germany), 200 µM of each dNTP (Boehringer Mannheim, Germany), 5 µL 10× PCR reaction buffer (Boehringer Mannheim, Germany), 50 ng of genomic DNA and 10 pM of each primer. PCRs were carried out using an MJ Research PTC-200 thermal cycler (MJ Research Inc., USA). Direct sequencing was performed using Big dye Terminator Kits (Applied Biosystems, USA) on an ABI PRISM 3100 DNA Sequencer (Applied Biosystems, USA). The single base extension method (SNaP shotR) was used to confirm SNP candidates obtained by sequence alignments of PCR products.
Bayesian estimate
We took peripheral blood samples of patients at 4 h after oral administration of the last dose when they attained steady-state after administrating theophylline at least 5 times. Since all blood samples were not collected at the same time after the last dose, it was necessary to extrapolate serum theophylline concentrations to a single time-point (12 h after the last dose) to facilitate comparisons. The pharmacokinetic software ABBOTTBASE pharmacokinetic systems v1.10 (Abbott laboratories, USA) was used to perform this extrapolation. Dosing regimens, measured theophylline concentrations in individual patients, and the following population pharmacokinetic-parameters were used to determined clearance values; volume of distribution (Vd) = 0.45 ± 0.10, bioavailability (F) = 1.00 (for Theoclear® and Etheophyl®), and absorption rate constant (Ka) = 0.50 or 0.40 (for Theoclear® or Etheophyl®).
Nuclear extract
Crude nuclear extracts for electrophoretic mobility shift assay (EMSA) were prepared by modifying a previously published protocol [23]. Briefly, the human hepatocyte cell line, B16A2, was used for nuclear extract preparation. Cells were grown in a 100 mm plate to 100% confluence and maintained for an additional 3-4 weeks before nuclear extracts were prepared. Cells were washed with ice-cold PBS twice, collected in 1ml PBS, and centrifuged at 3,000 rpm for 10 min at 4℃. Cell pellets were washed twice with PBS and resuspended in 1 mL hypotonic buffer A (10 mM HEPES, PH 7.9, 1.5 mM MgCl2, 10 nM KCl, 0.2 mM PMSF, 0.5 mM DTT, 10 µg/mL leupeptin), kept on ice for 10 min, and then centrifuged for 10 min at 4℃. Nuclear pellets were resuspended in 1 to 1.5 volumes of ice-cold salt buffer B (20 mM HEPES, PH 7.9, 1.5 mM MgCl2, 25% glycerol, 0.2 mM EDTA, 0.5 mM DTT, 0.5 mM PMSF, 10 µg/mL leupeptin), kept on ice for 30 min, and centrifuged at 15,000 rpm for 30 min at 4℃. The supernatant obtained was added to buffer C (20 mM HEPES, pH 7.9, 20% glycerol, 100 mM KCl, 0.2 mM EDTA with 1 mM phenylmethylsulfonyl fluoride (PMSF), 1 mM DTT) and dialyzed. Finally, nuclear protein concentrations were measured using the Bradford method [24].
EMSA (Gel retardation assay)
Double-stranded 12mer nucleotides were used as described previously [20]. These probes were phosphorylated with T4 polynucleotide kinase in the presence of [γ-32P] dATP. Gel retardation experiments were carried out by incubating labeled probes (5 fmol) with 6 or 12 µg of nuclear protein extract prepared from human liver in a buffer containing 20 mM HEPES, PH 7.9, 40 mM KCl, 0.5 mM DTT (Dithiothreitol), 0.1 mM EDTA, 1 mM MgCl2, 4% ficoll, 2 µg poly (dl-dC) at room temperature for 15 min, as described by Gorski et al. [25]. Samples were loaded onto a native polyacrylamide gel, and electrophoresis was carried out in 1xEPB (6.7 mM Tris, 1 mM EDTA, 3.3 mM sodium acetate, pH 7.5) at 130 V for 4 h. Unlabeled double-stranded oligonucleotides were added to incubation mixtures as competitors.
Statistical analysis
The nonlinear mixed effects model (NONMEM) program was used to determine mean theophylline clearance which is especially useful when there are only a few pharmacokinetic measurements from each individual sampled in the population, or when the data collection design varies considerably between these individuals [26].
The statistical significances of associations between polymorphisms of the CYP1A2 or CYP2E1 gene and theophylline clearance were assessed by analysis of variance (one-way fractional ANOVA) and the student's t-test. Also, multiple regression analysis was performed to assume relationship between theophylline clearance, genotype, age, severity, gender and atopy. p values of <0.05 were considered to indicate statistical significance. Statistical analyses were performed using SPSS for windows version 12.0 (SPSS Inc, USA).
RESULTS
Detection of genetic polymorphisms in the CYP1A2 and CYP2E1 genes
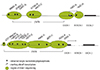
Amplified regions are shown (Fig. 1), and PCR products were obtained using the primers shown in table 2. Seven variants were detected, five in the 5' flanking and promoter regions (SNPs: -3860G>A, -3598G>T, -3594T>G, -3113G>A, -2847T>C) and two in intron 1 (-739T>G, -163C>A) of the CYP1A2 gene by direct sequencing (numbering according to GenBank Accession number NM_00761). In the CYP2E1 gene, ten polymorphisms were detected, though most polymorphisms were rare (<1%) except for -1053C>T and -1293G>C and a 96 bp insertion at -2257 (-1566T>A, -1515T>G, -1027T>C, -930G>G, -807T>C,-352G>A, -333A>T). The genotype distributions of these SNPs are shown in table 3 (numbering according to GenBank Accession number NT_017795).
Relationship between polymorphisms of the CYP1A2 gene and theophylline metabolism in asthmatic patients
Mean theophylline clearances were assessed according to genotypes in CYP1A2, in which significant association was observed at the -3860G>A point mutation. Those with an A mutant allele genotype (GA+AA) had significantly higher theophylline clearance than those with the wild GG genotype. We found a significant increase in the theophylline clearance in those with A mutant alleles versus those with the wild genotype (29.11 ± 0.91 mL/kg/h vs. 26.12 ± 0.80 mL/kg/h, p = 0.014, Fig. 2).
No association was found between theophylline clearance and the other six SNPs in CYP1A2 (p > 0.05) including -3598G>T which excluded from the regression model because the SNP was not appropriate for fitting the model (Table 4).
Contribution of the CYP2E1 gene and of various factors to theophylline metabolism in asthmatic patients
We preformed association analysis to examine the relations between theophylline clearance and the three SNPs in the CYP2E1. In addition, we also searched for relationships between theophylline clearance and other factors, i.e., age, gender, severity of asthma, and type of asthma, which are phenotypes known to affect theophylline clearance. But no significant association was found (p > 0.05, Table 4).
We evaluated that there were no gene-gene interactions of the genotypes evaluated in CYP1A2 and CYP2E1 by chi-square test between SNPs found in each gene (data not shown).
EMSA (Gel retardation assay)
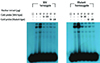
Gel retardation assays were performed using two types of DNA oligomers (GG and AA) presenting polymorphism at -3860 site of the CYP1A2 gene (Wild-type: 5'-GCC TCT CGG ATT-3', Mutanttype: 5'-GCC TCT CAG ATT-3') to determine whether any factor influenced the binding affinity of the variant at -3860G>A of the CYP1A2 gene. The results shown in figure 3 demonstrate that one protein bound to both types of oligomers. The formation of a complex with wild type oligomer was inhibited in the presence of 50- and 100-fold molar excesses of unlabeled wild type and mutant type competitor. The formation of a complex with mutant type oligomer was inhibited in the same way. Cross reactivity between wild type and mutant type fragments existed. However, the binding affinities of polymorphisms at the -3860 site of the CYP1A2 gene differed, whereby the binding affinity of the oligomer with the A allele was higher than that to the G allele at the -3860G>A point mutation.
DISCUSSION
Theophylline is widely used in asthma because of its effectiveness as an add-on treatment [27, 28] as well as safe for the elderly with asthma [29]. It is known to be metabolized predominantly by CYP1A2 and CYP2E1 with considerable inter-individual variability [10, 15]. Here we undertook to evaluate the association between genetic variants of CYP1A2 and CYP2E1 and the metabolism of theophylline in non-smoking Korean asthmatics.
Our study provides direct evidence of a relationship between theophylline clearance and the CYP1A2 genotype. Mean theophylline clearance by non-smoking Korean asthmatics with mutant variants (GA + AA) at the -3860 site of the CYP1A2 gene was significantly higher than of those with the GG allele, and in gel retardation assays, we found one transcriptional factor with different binding affinities at the -3860G>A polymorphic site. This transcription factor binding affinity was found to higher for the mutant A allele at the -3860 site than for the wild type, which suggests that higher transcriptional activity of the mutant variant. As a result, point mutation in the 5'-flanking region in human CYP1A2 might influence CYP1A2 inducibility and activity. Allele frequencies at -3860G>A of CYP1A2 gene were found to be similar in Korean and Japanese asthma patients [19, 30]. However, the tendency between genotype and phenotype that we found were different from the previous studies. Obase et al. [19] reported that mutant variants at the -3860G>A polymorphism are associated with reduced theophylline clearance in Japanese non-smoking asthmatics. Moreover, age and severity of asthma were found to affect theophylline metabolism in Japanese smoking subjects. Nakajima et al. demonstrated that at least two factors bind to wild and mutated oligonucleotides in Japanese subjects by using EMSA [20]. This finding is quite different from that of the present study, which demonstrates that one transcriptional factor has different binding affinity at the -3860G>A polymorphic site in Korean non-smoking asthmatics. This result indicates the possibility of an ethnic difference between Korean and Japanese non-smoking asthmatics in terms of liver CYP1A2 transcriptional activity.
Recently, Yoon et al. [30] also concluded that CYP1A2 and CYP2E1 polymorphisms are associated with theophylline metabolism in Korean asthmatics. They showed that mutant variants at the -3860G>A polymorphism are associated with reduced theophylline clearance. Although it's hard to conclude which factors influenced on the difference between those studies, because the previous study didn't cited whether they control the affecting factors in CYP1A2 as well as CYP2E1 when they enrolled the subjects, differences between the previous study and our study could have been caused by several factors, e.g., the co-administration of drugs or smoking habit or the severity of asthma [25], all of which can influence theophylline clearance.
And the methods to determine mean clearance value could cause the difference among those studies, which we determined it by using population PK-parameters. Theophylline is N-demethylated to 3MX and 1MX and is converted into 1,3-DMU via 8-hydroxylation which accounts for about half of its clearance when orally administered [4]. The other studies could account for that the oxidation of theophylline might be related to CYP1A2 or CYP2E1 polymorphisms [19, 30] because they used ratio of 1,3-DMU/theophylline to determine the clearance value, however, that is a partial information of metabolic pathway of theophylline accounting for only 35-45% of the drug recovered in urine [31], not included demethylation pathway. To evaluate theophylline clearance value using population PK data is common by NONMEM which is a general program and can be used to fit models to a wide variety of data and this study is valuable for connecting information between SNPs of CYP1A2 and patients' clearance data which is estimated by volume of distribution and absorption rate constant (Ka) for theophylline dosage form using population PK data. Previous studies have reported that different correlations between phenotype and genotypes, -3860G>A of CYP1A2, among the same genetic back ground in Japan, that is similar to this study [32]. And another studies have produced different results concerning genotypes at same sites and relationships between these and phenotypes [33, 34], which has led several groups to suggest that environmental factors should be explored in addition to heredity [14, 35]. For example, Tantcheva-Poór et al. [35] and Ou-Yang et al. [36] in studies on Chinese populations, showed that the activity of CYP1A2 in the Qidong population was significantly higher than in the Changsha population (who have a lower risk hepatocellular carcinoma). In fact, the mean clearance of theophylline in patients with the GG genotype at the -3860G>A site was much higher in the Obase group than in our group (34 ± 0.1 mL/kg/h vs. 27.53 ± 0.62 mL/kg/h) [19]. This difference may have been due to environmental characteristics that affect theophylline clearance through the induction of transcription factors and an increased hepatic function. Thus, we suggest that there may be significant environmental differences between Japanese and Korean subjects.
The frequency of SNPs found in CYP2E1 gene was evaluated. There was no difference between the frequency of the variants in patients and that of the matched 50 controls (p = 0.45) except for 3 SNPs including 96 bp insertion at -2257 and -1053C>T and -1293G>C which is known to affect theophylline clearance. We found -1053C>T and -1293G>C of CYP2E1 was in complete linkage disequilibrium as previous studies [12, 37]. However, there was not complete linkage disequilibrium of 5 SNPs which Yoon's group reported [30]. And -807T>C of CYP2E1 showed incomplete linkage disequilibrium. There reported several factors which causes statistically significant differences observed in allele frequency among study groups in CYP2E1, in relation to ethnicity, gender, sample size which causes type I statistical error, methods to confirm the allele frequency [38-40].
The discrepancy of allele frequency between present and previous study might come from following factors. First, small sample size of both studies could be the causal factor of the difference. Above mentioned, there was no difference between the frequency of the variants in patients and that of the matched 50 controls, however the allelic frequencies of wild homozygote at 96 bp insertion and -1053C>T and -1293G>C in CYP2E1 tends to reduce if sample size is reduced (96 bp insertion: 79.3% (n = 323) [41] vs. 71.9% (n = 89) vs. 64.5% (n = 62) [30]; -1053C>T and -1293G>C: 77.2% (n = 323) [41] vs. 66.3% (n = 89) vs. 62.9% (n = 62) [30]).
The differences between those studies were not only for the frequency but also for the expected enzyme activity of the same genotypes. There were some controversial studies that showed different tendency between genotype and phenotype for the rare mutant allele of CYP2E1, which is associated with higher transcriptional activity than the more common wild-type allele [12, 37]. However, the other studies do not show any relationship between the polymorphism and the enzyme activity [42, 43]. Therefore, it is necessary to come up with functional study if there showed significant relationship between genotype and phenotype to confirm the result authentically. We assessed the relationship between those SNPs known to affecting phenotype and theophylline clearance (one-way ANOVA) and found that there was no relationship between genotypes in the 5'-flanking region of the CYP2E1 gene and theophylline clearance or in the gene-gene interactions of the genotypes evaluated in CYP1A2 and CYP2E1 by chi-square t-test among SNPs found in those gene. We surmise that CYP2E1 polymorphism do not affect 8-hydroxylation of theophylline based on an association study of SNPs and theophylline clearance in non-smoking Korean asthmatics.
In Conclusion, we examined the associations of genetic polymorphisms among CYP1A2 and/or CYP2E1 and performed functional study with a polymorphism of -3860G>A in CYP1A2 gene, which showed the association with enzyme inducibility that have not been investigated simultaneously in asthmatic patients. The results of the present study suggest that increased theophylline clearance was significantly related to the -3860G>A polymorphism, and this polymorphism could influence the enzyme inducibility of CYP1A2 in Korean non-smoking asthmatics. Our study may be useful in terms of providing pharmacogenetic information for optimal use of theophylline to get the best treatment effectiveness.





 XML Download
XML Download