

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
There is a dire need to understand the underlying pathophysiology of asthma, including the complex genetic and environmental influences, to develop more effective treatment strategies. Though the role of respiratory pathogens in asthma has been extensively studied over the past few decades, many questions remain unanswered. First, of the known pathogens, which are the most likely to cause an infection associated with recurrent wheezing/asthma development? Second, do early childhood infections with respiratory pathogens cause asthma or do they merely select those that are predisposed to these infections? Third, if they are a causal factor, what is the immunopathology behind it? In the following sections, the interactions between respiratory infections and asthma development/exacerbations will be explored, with an emphasis on those pathogens associated with eosinophilia.
Infections in infancy and asthma development viral infection
It is well known that a substantial portion of infants who require hospitalization for bronchiolitis develop asthma by the age of 13 [1-5]. The importance of early life infection has been highlighted by studies showing the timing of birth in relationship to the winter virus season can increase the risk of developing asthma by age 6 years by as much as 30% [6]. The exact mechanisms and pathophysiologic pathways by which infection leads to asthma are not known, and those that have been proposed are subject to intense debate. Risk factors for bronchiolitis include young age (especially <6 months), small lung size and exposure to tobacco smoke [7]. Despite the uncertainty, it is generally considered respiratory infection does not work alone, but rather synergistically with respiratory allergy to produce immunologic and physiologic conditions conducive to asthma development.
Respiratory syncytial virus (RSV) has been the main focus of research, and most infants will have had at least one infection with RSV by the age of three. This virus is the major cause of bronchiolitis in children <1 year of age; consequently, it is considered the most important respiratory tract pathogen of early childhood [8]. No direct interventional studies demonstrating a causal relationship between RSV and asthma have been published to date. However, a long-term study of over 95,000 children found that timing of birth in relationship to the winter virus peak (particularly RSV) independently predicted asthma development, with the highest risk estimated for those born 121 days (~4 months) before the peak [6]. In addition, the administration of anti-RSV immune globulin (palivizumab) to children at high risk of chronic airway disease improved lung function and reduced the incidence of asthma and atopy, suggesting the prevention of RSV infection has long-term effects on respiratory and immunologic parameters relevant to asthma development [9]. In a larger more recent study, palivizumab significantly reduced recurrent wheezing [10]. However, because nearly every child has been infected with RSV at least once, other genetic, environmental, and/or developmental factors must also contribute to the epidemiological link with childhood asthma [11].
A number of recent papers have highlighted human rhinovirus's (HRV) possible role in asthma etiology. HRV is now recognized as an important cause of wheezing illness; however, it is most often found in asymptomatic infants and children [12, 13]. The wide variety of illness severity may be due to either host factors (abnormal innate immune responses) [14] or virus strain (more virulent or pathogenic strains) [15]. In comparative studies, children with RV-induced bronchiolitis were older and more atopic [16], and had a more severe disease course in the acute phase [17] than children with RSV-induced bronchiolitis. In the Perth Birth Cohort study, HRV was the most common pathogen associated with an acute respiratory infection in the first year of life followed by RSV [18]. Another study found wheezing HRV illnesses during the first 3 years of life were associated with a nearly 10-fold increase in asthma risk at age 6 years [19]. In contrast, several studies suggest HRV infection is not a risk factor for asthma, but may instead reveal children predisposed to asthma due to abnormal lung physiology and/or immune responses [18, 20, 21].
Another virus that has been implicated as a possible cause of wheezing and asthma is human bocavirus (HBoV), a novel parvovirus first isolated in 2005 from the respiratory secretions of patients with pneumonia [22]. However, the link between HBoV and asthma, or even any respiratory disease, has been complicated by the fact that it has a high rate of co-infection [23]. A recent study conducted at Inje University Sanggye Paik Hospital, found an association between HBoV infection and acute wheezing in children [24]. In their epidemiological study on acute wheezing and children, HBoV (13.8%) was the third most frequently found virus after RV (33.3%) and RSV (13.8%). Another epidemiological study by Vallet et al. [25] found HBoV infection to be associated with 13% of severe asthma exacerbations in children. They suggested HBoV could play a major role in acute exacerbations in asthmatic children.
Other viral infections during infancy and early childhood causing lower respiratory infections are also associated with recurrent wheezing and asthma development. These include parainfluenza, influenza A and human metapneumovirus (HMPV) [20, 26, 27]. In our recent study, we showed HMPV infection was associated with recurrent wheezing in children [28].
Bacterial infection
The role of bacterial infection in the pathogenesis of acute and chronic asthma remains quite controversial [29], and of the bacterial respiratory pathogens, the atypical bacteria Mycoplasma (M.) pneumoniae and Chlamydophila (C.) pneumoniae are the most commonly implicated. A 2011 study of 407 children under 5 years of age found M. pneumoniae and C. pneumoniae in approximately 10% and 2% of acute respiratory tract infections, with M. pneumoniae being associated with more severe disease including hospitalization for bronchiolitis [30]. In a follow-up study of 50 children with a previous history of M. pneumoniae, 23 suffered from recurrent wheezy bronchitis or asthma, including 5 who developed wheeze for the first time [31]. Case reports of chronic asthma beginning with M. pneumoniae infection also suggest this pathogen is a potential causative agent of asthma [32].
It has been suggested that chronic bacterial infection or colonization with pathogenic bacteria could eventually lead to asthma. In Bisgaard et al.'s study [33], they found that neonates colonized in the hypopharyngeal region with Streptococcus pneumoniae, Haemophilus influenzae or Moraxella catarrhalis (or a combination of them), but not Staphylococcus (S.) aureus, were at increased risk for recurrent wheeze and asthma by 5 years of age. However, other studies have shown an association between S. aureus and atopic diseases such as dermatitis, rhinitis and asthma [34-36]. More specifically, staphylococcal enterotoxin exposure has been associated with the pathogenesis of asthma and/or atopic dermatitis [36-39]. A high prevalence of C. pneumoniae infection in school-age children with wheezing has also been found [40].
The mounting data detailing the strong association between respiratory viral/bacterial infection and asthma suggests further studies are needed to clearly demonstrate causality. An alternative mechanism may be that individuals with a tendency toward atopy may have increased susceptibility to bacterial and viral infections and an increased risk of asthma development [16, 20, 41, 42]. The question of whether early childhood infections cause asthma, or instead cause chronic disease in those with an underlying predisposition to asthma, remains unanswered.
Infection and asthma exacerbations
It has been estimated that nearly 85% of asthma exacerbations in children, and nearly 50% in adults, are a result of viral infections [43, 44]. Indeed, with the advent of nucleic acid amplification testing (e.g., reverse transcription-polymerase chain reaction) and immunoassays, the detection of HRV and other RNA viruses has been enhanced suggesting HRV may be responsible for a larger proportion of exacerbations than previously thought. In a study by Rakes et al. [45], HRV was the predominant pathogen (71%) in children aged 2-16 presenting to emergency departments with acute wheezing, while only 6% were positive for RSV. A study of 206 asthmatic children aged 3-18 found HRV (26%) as the most common cause of exacerbation [46], while in adults, HRV was again the most common pathogen (56% of virus-positive specimens) found in asthma-related acute-care visits [47]. These studies provide strong evidence of the association between respiratory viral infection and asthma exacerbation. However, it is still unclear whether viral infection alone can exacerbate asthma, or if other factors, such as exposure to allergens [48] or air pollution [49], work synergistically to precipitate an attack.
Several studies have also demonstrated an association between atypical bacterial infection and asthma exacerbation. C. pneumoniae has been implicated in severe chronic asthma [50, 51] and both C. pneumoniae and M. pneumoniae have been linked to acute asthma exacerbation in children and adults [52, 53]. In a recent study [54], 22 of 58 (39%) patients presenting with acute exacerbation of bronchial asthma were found to be infected with either C. pneumoniae, M. pneumoniae or both, and those with atypical infection had greater functional impairment on admission when compared to patients with non-atypical respiratory infection.
The body's response to an invading pathogen includes the respiratory, immune and nervous systems [55]. Respiratory infections during infancy may have acute and long-term effects on lung and immune system development, and represent a risk factor for asthma development. When bronchial epithelial cells are infected by a pathogen (Fig. 1), they generate both local and systemic immune responses aimed at clearing the infection. Neural signals are also generated to coordinate inflammation. In the asthmatic individual, however, immune response is altered and bronchial epithelial cells are damaged. A damaged bronchial epithelial lining is more susceptible to penetration by environmental irritants, which can directly stimulate sensory nerves leading to increased histamine release from mast cells and induction of smooth muscle cell contraction (i.e., bronchoconstriction) by released kinins [56]. A number of mediators are released by infected bronchial epithelial cells and play a role in recruiting major inflammatory cells, such as eosinophils, neutrophils and mast cells, though these cells may already be present in asthmatic airways. Eosinophils can be recruited to inflammatory sites by cytokines (most notably the T-helper type 2 (Th2) cell-derived IL-4, IL-5, and IL-13), and chemokines CCL5/RANTES and CCL11/eotaxin-1 [57], while neutrophils are most likely recruited by CXCL8/IL-8 or LTB4 [58]. Mast cells can, in turn, be activated by eosinophil-derived major basic protein (MBP) to release a number of mediators [59]. Together, these major inflammatory cells release mediators that can cause airway hyperresponsiveness (AHR), airway remodeling (through the action of eosinophils, fibrogenic and growth factors), and airway limitation (bronchoconstriction, mucosal edema, hypersecretion), leading to characteristic asthma exacerbation symptoms (wheeze, dyspnea, cough, chest tightness).
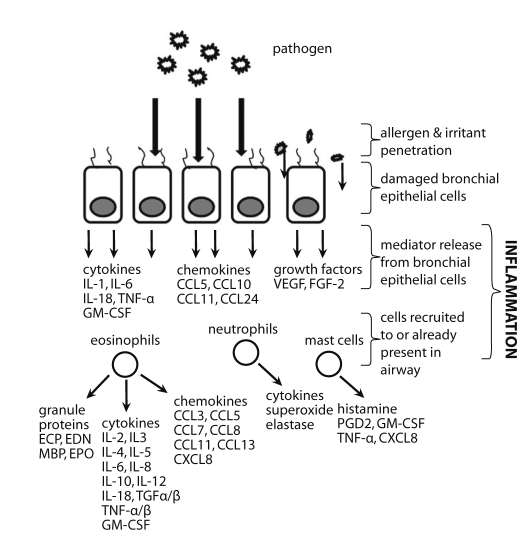 | Fig. 1Infection of bronchial epithelial cells leads to release of mediators, including cytokines, chemokines, and growth factors. Recruited immune cells (eosinophils, neutrophils, and mast cells) release a number of mediators, as well. CCL, CC chemokine ligand; CXCL, CXC chemokine ligand; ECP, eosinophil cationic protein; EDN, eosinophilderived neurotoxin; EPO, eosinophil peroxidase; FGF-2, fibroblast growth factor-2; GM-CSF, granulocyte macrophage-colony stimulating factor; IL, interleukin; LT, leukotriene; MBP, major basic protein; PAF, platelet activating factor; PG, prostaglandin; TNF, tumour necrosis factor; VEGF, vascular endothelial growth factor.
|
Eosinophils are generally considered as major effector cells of asthma, and it has long been thought that the eosinophilic response to viral infection has a predominantly negative effect on human health, that it is the major cause of inflammation responsible for tissue damage, bronchoconstriction and respiratory dysfunction [60]. However, several studies have shown eosinophils may promote viral clearance and antiviral host defense. This concept, termed the "double-edged sword", was first introduced by Smith [61] in 1994, with respect to the dual role of neutrophils in viral infection. Rosenberg and Domachowske [62-64] followed with their hypothesis that eosinophils may be recruited in part to promote primary antiviral host defense, possibly in situations where acquired immune response was less than effective [65]. More specifically, through their secretory mediators, eosinophils could reduce the infectivity of RSV for target epithelial cells in vitro. More recently, Phipps et al. [66] demonstrated accelerated clearance of RSV from murine lungs, and that full antiviral activity was dependent on intact TLR signaling in eosinophils introduced exogenously.
The clinical course of Mycoplasma pneumonia is typically mild and self-limited; however, long-term complications can occur via three possible mechanisms. According to a study by Kim et al. [67], 14 of 38 (37%) children hospitalized with Mycoplasma pneumonia had abnormal high-resolution computed tomography findings (i.e., mosaic perfusion, bronchiectasis, bronchial wall thickening, air trapping, and Swyer-James Syndrome) suggestive of small airway obstruction. Long-term complications may also occur due to immune modulation. Koh et al. [68] found increased IL-4 levels and IL-4/interferon (IFN)-γ ratios in patients with Mycoplasma pneumonia. The data suggested a predominant Th2-like cytokine response, thus representing a favorable condition for IgE production that may result in the release of chemical mediators leading to bronchospasm, airway inflammation, and airway hyperresponsiveness. A third possible mechanism by which M. pneumoniae infection can lead to long-term complications is through functional abnormalities. Pulmonary function test results, such as decreased flow rate at 50% of vital capacity [39], have been reported. Furthermore, M. pneumoniae infection has been associated with bronchial hyperresponsiveness [69]. Long-term impairment of small airways function has also been noted in studies by Mok et al. [31], Sabato et al. [70] and several others.
Infection and the hygiene hypothesis
Early childhood exposure to certain pathogens may actually be protective against atopy and/or asthma, termed the "hygiene hypothesis" [71]. This highly controversial idea was borne from early observations showing the risk of developing allergy and/or asthma was inversely related to the number of children in the family [72]. However, this seems paradoxical, as it is also known that bronchiolitis and pneumonia in infancy lead to an increased risk of subsequent asthma. Whether infections have a protective effect or not may have to do with location, frequency, intensity, and timing [34, 73, 74].
Another possible mechanism for the switch to an atopic phenotype has been proposed; namely, a defect in the stimulation of dendritic cells by nonpathogenic microorganisms in gut-associated lymphoid tissue leading to reduced production of regulatory T cells [75, 76]. Differences in gut bacteria colonization have been reported in children with atopic dermatitis. For example, Enterococci and Bifidobacteria were reduced, while Clostridia and S. aureus were increased [77, 78]. These changes have been partly associated with the increased use of antibiotics [34].
Other environmental factors influence the development of atopic sensitization, including early exposure to pets and an agricultural background [79]. In an agricultural setting, increased exposure to high levels of endotoxin has been associated with lower rates of allergy and an elevated IFN-producing cells in the blood [80, 81], cells which are known for their potent antiviral activity [82]. Taken one step further, Schaub et al. [83] demonstrated maternal exposure to farming decreases the risk of allergic disease in offspring, possibly through the increased number and activation of T-regulatory cord blood cells associated with lower Th2 cytokine secretion and lymphocyte proliferation on innate exposure. Indeed, there is growing evidence, both in human and animal studies, that pre- and post-natal exposure to pathogens and allergens may provide a protective effect against allergy development. In addition, breastfeeding has been shown to protect against asthma and lower respiratory illness, especially RSV [84, 85].
Infection and atopy
One paramount question that remains unanswered in determining the link between early childhood infection and asthma development is: Do respiratory infections merely select those individuals that are predisposed to asthma, or are these infections able to alter lung development/immune response enough to actually cause asthma? A number of studies have shown that respiratory allergy may play a potentially synergistic role with viral/bacterial infections to produce airway inflammation and subsequent asthma in childhood [20, 73, 84]. One recent cohort study by Kusel et al. [20] found only an association between viral infections during infancy and subsequent development of persistent wheeze and asthma at 5 years in children with atopic sensitization during the first 2 years of life. This association was also restricted to infections that spread to the lower respiratory and were intense enough to cause severe symptoms in the infants. Together, respiratory allergy and infection can cause airway dysfunction through several mechanisms, including: viral infection damaging barrier function of the airway epithelium, leading to enhanced absorption of aeroallergens [86], and the generation of various cytokines, chemokines, leukotrienes, and molecules that may further upregulate cellular recruitment, cell activation, and the continuing inflammatory response [56].
The role of eosinophilia in asthma development/exacerbation
Eosinophilic inflammation is a cardinal feature of asthma and increases in eosinophilic inflammatory markers have been shown to be good predictors of asthma exacerbations [87, 88]. It has been demonstrated that suppression of eosinophilic inflammation with glucocorticosteroids is associated with an amelioration of symptoms and airway dysfunction [89]. A recent study by Fanat et al. [90] found anti-IL-5 treatment reduced the eosinophilopoietic potential of airway smooth muscle cells, suggesting it may promote in situ eosinophilopoiesis in asthmatic lungs. Furthermore, Haldar et al. [91] and Nair et al. [92] demonstrated the ability of anti-IL-5 therapy in reducing eosinophil numbers while improving asthma control in refractory eosinophilic asthma.
Eosinophils as effector cells
Eosinophils are major effector cells of the allergic process. Activation of eosinophils leads to extracellular release of a number of granule proteins, such as eosinophil cationic protein (ECP), eosinophil-derived neurotoxin (EDN), MBP, and eosinophil peroxidase (refer to Fig. 1) [93]. Among the many mediators released during eosinophil activation and degranulation, it is these eosinophil granule proteins that are the most strongly implicated in the pathophysiology of asthma [94]. Eosinophils also release a number of proinflammatory cytokines, chemokines and lipid mediators. In addition, eosinophils express a large number of cell-surface markers, including adhesion and apoptotic signaling molecules, chemokine, complement and chemotactic factor receptors, and cytokine, immunoglobulin, prostaglandin, platelet activating factor, and leukotriene receptors [95-99].
Eosinophils have generally been regarded as terminal effector cells in allergic airway diseases; however, recent studies demonstrate their involvement in the initial stages of allergic disease development, as well [100]. Murine studies suggest eosinophils may actually drive T-cell responses as opposed to merely being driven by them. Eosinophils can act as antigen-presenting cells [101, 102]; thus, at disease onset they are able to induce activation and proliferation and/or cytokine production in T cells.
Viral infection and eosinophilia
To further elucidate the connection between early childhood viral infection and asthma development, several studies involving RSV bronchiolitis have been carried out due to the clinical analogy, epidemiologic relationship, and similar immunologic phenomena in the airways, such as specific IgE production [103], generation of chemokines [104] and adhesion molecules [105], and eosinophil activation [106] it shares with childhood asthma. A number of studies have demonstrated eosinophilia during/post RSV bronchiolitis [16, 45, 106-110]. However, the downfall of many RSV studies is the lack of premorbid eosinophil levels. Therefore, it cannot be explicitly stated that the observed eosinophilia was a result of RSV infection. Indeed, prenatal Th2/Th1 imbalance, allergic sensitization during pregnancy or some other reason may result in elevated eosinophil levels in infants. This was suggested in a study by Frischer et al. [111] of neonates born to atopic parents.
Though most viral infections are thought to be neutrophilic in nature, studies by our research group have shown a subgroup of RSV bronchiolitis patients is eosinophil-positive [107]. This subgroup also had higher levels of Th2 cytokines. Eosinophilia together with a skewed Th2 cytokine response strongly resembles the immunologic profile of childhood asthma. To further strengthen the relationship between RSV bronchiolitis and asthma, our research group recently undertook a RSV bronchiolitis study [106] investigating eosinophil degranulation post-bronchiolitis and its association with recurrent wheezing. We found good correlations between the eosinophil degranulation marker, EDN, and recurrent wheezing episodes in a cohort of infants with a first episode of RSV bronchiolitis. Furthermore, EDN levels at 3 months proved to have predictive value for recurrent wheezing (positive predictive value = 57%, negative predictive value = 76%, sensitivity = 72%, specificity = 62%). Indeed, it appears there is a subgroup of RSV bronchiolitis patients with marked eosinophilia that may go on to later develop asthma. This hypothesis may help elucidate the link between early childhood infections and asthma development.
Bacterial infection and eosinophilia
One connection between bacterial infection and asthma pathogenesis may be the eosinophil. An in vivo study found patients with acute bacterial infection exhibited increased levels of serum ECP levels [112]. The authors concluded that eosinophils are activated during acute bacterial infections and that this activation results in the preferential release of ECP. Another study [113] found elevated ECP levels (>200 ng/mL) in all 16 patients presenting with acute community-acquired bacterial sinusitis, and 10 of those 16 had remarkably elevated ECP levels (>3,000 ng/mL). It should be noted that these patients had no history of chronic sinusitis, active or chronic allergies, or any other chronic upper respiratory disease. An in vitro study found pathogenic bacteria induced EDN release by eosinophils [114], and in a murine model, staphylococcal enterotoxin B induced eosinophilic airway inflammation and evidence of eosinophil degranulation [115].
Elevated levels of ECP have been associated with the increased activation of eosinophils [116]; hence, ECP is thought to reflect the activation state of eosinophils [117]. The immunologic benefits of eosinophil activation and ECP release may include antibacterial activity in concentrations 10-20 µg/mL and above [118, 119]. It has been speculated that due to its stability, ECP may accumulate over time in mucus fluids and act as a first line of defense against bacterial invasion [120].
Asthma phenotypes
A number of techniques have recently been used to characterize the myriad of asthma phenotypes in hopes of better tailoring treatment to the individual. The classical IgE-associated allergic asthma phenotype (also termed 'extrinsic') starting in childhood is the most widely studied, as it is the most common in real-life and easily studied in the clinical laboratory. In allergic asthma, the cytokine profile is predominantly Th2 (i.e., IL-4, IL-5, IL-9, and IL-13). A recent genome microarray study by Woodruff et al. [121] found significant IL-13-mediated gene induction in asthma. One genetic biomarker in particular was associated with eosinophil accumulation in the airways, while another was identified as a potential mediator of corticosteroid-resistant asthma. Other studies, including the one by Martin et al. [122], have vetted other predictive markers, such as IgE and blood eosinophil levels, serum eosinophil degranulation protein levels, lung function, and PC20 [123]. These examples highlight the need for careful consideration of phenotypes, especially variable treatment responses, when treating asthma patients.
The less common 'intrinsic' asthma is of late-onset, lacks circulating specific IgE, and no sensitivities to allergens can be identified [57]; despite this, airway eosinophilia and Th2 cells feature prominently [124]. Intrinsic asthma is less common (~10%) than extrinsic asthma, is more common in females, and tends to be more severe, requiring higher doses of corticosteroids [125]. Clinically, however, intrinsic asthma shares a great deal with extrinsic asthma, including variable airflow obstruction and symptoms, and a good therapeutic response to corticosteroid therapy. It is well established that upper respiratory tract viral infections are the most common cause of exacerbations in intrinsic and extrinsic asthma [126].
In addition to eosinophilic inflammation, other cardinal features of most asthmatic phenotypes include AHR, excessive airway mucus production, and airway remodeling [57].
Eosinophils and asthma
Eosinophil increases in the tissues, body fluids (e.g., sputum, peripheral blood, serum, bronchoalveolar lavage) and bone marrow have been noted in asthma [107, 127, 128]. These elevated levels have correlated well with disease severity, leading to the hypothesis that the eosinophil is the major effector cell of asthma. One way in which they act as major effector cells is through the release of granule proteins, which induce tissue damage and dysfunction [129], as well as the further propagation of airway inflammation. Elevated levels of ECP [130] and EDN [131] have been noted in asthmatics, with higher levels of ECP [132] and EDN [133] found during asthma exacerbation when compared to healthy patients and those with stable asthma. This would suggest airway inflammation associated with asthma exacerbation is characterized not only by an increase in number of eosinophils, but also an increase in airway eosinophil degranulation [134].
Monitoring of asthma with eosinophil markers
Though eosinophils are important in the pathophysiology of asthma, eosinophil degranulation and its associated products, such as EDN and ECP, may be even more important. It has been suggested that the secretory activity of eosinophils - a combination of concentration of eosinophils and their propensity to release degranulation products - may be a key marker of disease activity and is more accurately measured by eosinophil degranulation products such as EDN and ECP [135]. Kim et al. [136] found significant differences in EDN and ECP levels in asthmatics during both acute and stable phases when compared to controls. Thus, EDN, along with ECP, may aid in the diagnosis of asthma. It has been suggested by several groups that EDN is more useful than ECP in evaluating disease severity [136-138]. This may partially be due to EDN's recoverability, as ECP is a sticky and more highly charged protein [129].
In terms of clinical utility, EDN levels are a more accurate biomarker of the underlying pathophysiology of asthma (i.e., eosinophilic inflammation); consequently, they provide an objective measure of the eosinophil's secretory activity. In children too young to fully participate in lung function tests, EDN levels may be useful as an alternative measurement of eosinophilic inflammation, but larger studies should be carried out to determine its reproducibility and repeatability for this purpose.
Go to : 
CONCLUSION
Viral and bacterial infections are important causes of childhood respiratory disease. They can lead to long-term morbidity, such as recurrent wheezing, and evidence points toward a very strong link between early childhood infection and asthma development. However, the underlying pathophysiologic link between infection and chronic allergic disease is still not fully understood. With increased understanding and attention to these disease processes, clinicians will better be able to diagnose, treat and monitor asthma. Indeed, focus must shift from not only treating symptoms, but also the underlying pathophysiology to provide optimal care to the patient.
Go to :
 XML Download
XML Download