

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Since the discovery of IgE there has been an explosive increase in knowledge about how IgE is generated and its role in driving mast cell, basophil and eosinophil mediated inflammatory responses underlying allergic disorders. Principal among these responses are the production of cytokines, chemokines and adhesion molecules associated with the Th2 profile, the release of a variety of preformed and newly generated mediators from mast cells, and the selective recruitment and activation of secondary effector cells, particularly eosinophils and basophils.
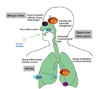
Allergic rhinitis (AR) is the most common of all allergic disease affecting approximately 400 million people worldwide and is associated with a reduced quality of life of the patients, lower work productivity and school learning performance as well as increasing medical costs [1-6]. Allergic rhinitis is an IgE-mediated inflammatory disease of the nasal mucous membranes due to the interaction of allergen characterized by an inflammatory infiltrate made up of eosinophils, T cells, mast cells and basophils, which release several mediators, chemokines and cytokines (among these, histamine and cysteinyl-leukotrienes are the major vasoactive mediators), regulation of the local and systemic IgE synthesis, and communication with the immune system and the bone marrow [7]. Clinically this results in the typical symptoms of rhinorrhea, sneezing, itching and nasal blockage. Systemic circulation of inflammatory cells allows their infiltration into other tissues where chemoattractant and adhesion molecules already exist. Thus, besides local inflammation, AR also triggers a systemic inflammation, which can in turn augment inflammation in both the upper and lower airways. Consequently, AR is linked to various comorbid conditions: asthma, rhinosinusitis, nasal polyposis and serous otitis media and sleep disorders [8, 9] (Fig. 1).
Genetics and epigenetics
Allergic rhinitis typically starts in early childhood with the symptoms usually peaking in the second, third, and fourth decades of life. However, an onset of this disease in infancy or later in adults is not quite uncommon. Like other atopic diseases, in allergic rhinitis multiple family members often are affected. Although genetic determinants of total IgE levels and specific allergen sensitization are suggested, neither the form of atopic illness nor specific sensitivities seem to be heritable as simple genetic traits. Although there are several studies of genes associated with asthma those for allergic rhinitis are rather few, limited to family genome wide association studies as well as single nucleotide polymorphisms. Furthermore, these studies have not been well replicated, had been done on small subject numbers, and often the subjects had co-morbid asthma or atopic dermatitis. An association with polymorphisms of the transcription factor GATA binding protein 3; trans-acting T-cell-specific transcription factor and interleukin (IL)-13 with the risk of developing allergic rhinitis was reported in a cohort of children followed up until the age of 10. Further studies are needed to identify candidate genes for allergic rhinitis.
There is increasing evidence pointing to the influence of prenatal and early life exposures on the development of allergic disease. Transient environmental pressures can have permanent effects on gene regulation and expression through epigenetic mechanisms. Histone modifications have been associated with degree of bronchial hyperresponsiveness and corticosteroid resistance in asthma. Epigenetic mechanisms can operate affect differentiation of human T cells and modify dendritic cells [10]. Murine studies have revealed that a maternal diet rich in methyl donors can enhance susceptibility to allergic inflammation in the offspring, mediated through increased DNA methylation [11]. Thus current evidence supports a role for epigenetics in the development and persistence of allergic rhinitis and asthma.
Early and late phase allergic reactions
After deposition and elution of allergens into the mucus layer, the allergens are taken up by antigen presenting cells and processed and presented to helper T lymphocytes. Activated helper T lymphocytes release cytokines like IL-4 and IL-13 and interact with B lymphocytes to induce the synthesis of allergen specific IgE. Thereafter, the allergen-specific IgE binds to the high affinity receptor for IgE on the surface of mast cells.
Early phase response
The early or immediate phase response occurs in sensitized individuals within min of exposure to the allergen and lasts for about 2-3 h. One of the cardinal components of the early phase response is the degranulation of mast cells. In the sensitized individual mast cells are abundant in the epithelial compartment of the nasal mucosa and can be easily activated upon re-exposure to the allergens. Upon crosslinking of the allergen specific IgE bound to the surface of mast cells by the specific allergen, mast cells degranulate and release a variety of pre-formed and newly formed mediators leading to what is known as the early phase response. Histamine, which is the major mediator of allergic rhinitis, stimulates the sensory nerve endings of the Vth nerve (trigeminal) and induces sneezing. Histamine also stimulates the mucous glands causing the secretion of mucous (rhinorrhea) and histamine, leukotrienes and prostaglandins acts on the blood vessels causing nasal congestion [7].
Late phase response
The early phase response is usually followed by the late phase response which occurs 4-6 h after antigen stimulation. The late phase response is characterized by a prolongation of symptoms - sneezing, rhinorrhea but most predominantly a sustained nasal congestion which lasts for about 18-24 h. The late phase response is predominantly inflammatory in nature and is characterized by a inflammatory cellular influx comprising of predominantly T lymphocytes, basophils and eosinophils [7]. A variety of mediators are released by these cells including leukotrienes, kinins, histamine which result in the continuation of the symptoms and the development of the late phase. The key to the orchestration of the late phase response lies in the production and release of a variety of cytokines and chemokines like IL-4, IL-13 from mast cells [12-14] as these cytokines can upregulate the expression of 'adhesion molecules' like vascular cell adhesion molecule 1 (VCAM-1) on the endothelial cells facilitating the infiltration of eosinophils, T lymphocytes and basophils into the nasal mucosa. In addition, chemokines like RANTES, eotaxin, MCP-4 and Thymus-and activation Regulated chemokine (TARC) released from epithelial cells serve as chemoattractants for eosinophils, basophils and T lymphocytes [15-18] (Fig. 2). Other cytokines like granulocytemacrophage colony stimulating factor (GM-CSF) released largely by epithelial cells and IL-5 from mast cells and T lymphocytes prolong the survival of the infiltrated eosinophils in the nasal mucosa [14]. More recently, mast cells have been found to further contribute to the late phase response through the histamine/tryptase-induced upregulation of GM-CSF and RANTES in nasal epithelial cells and the synergetic action of IL-4/IL-13 and TNF-α-inducing the upregulation of eotaxin and TARC production in nasal epithelial cells [19] (Fig. 2). In addition, a variety of other mediators released like eosinophil cationic protein (ECP), platelet activating factor, major basic protein (MBP) are also implicated in the late phase response.
Inflammatory cells and their regulation of allergic inflammation
Mast cells play a central role in the immune mechanisms of allergic rhinitis. Irani et al. [20] were the first to classify human mast cells into two phenotypically distinct subpopulations based on the type of neutral proteases they express, namely MC (T) that contain only tryptase, and MC (TC) that contain chymase, cathepsin G and carboxypeptidase in addition to tryptase. In patients with allergic rhinitis mast cells mast cells are found to abundantly accumulate in the epithelial compartment of the nasal mucosa [21] and these are predominantly of the MC (T) phenotype [21]. Recently the factors that induce the selective accumulation of mast cells into the allergic nasal epithelium has been of considerable interest to several researchers. Nillson et al. [22] have suggested stem cell factor (SCF) as the key molecule and Salib et al. [23] have suggested that TGF-β is implicated in this mast cell migration. We have shown that RANTES is the key molecule regulating the migration of mast cells into the allergic nasal epithelium [13] (Fig. 2).
In addition to the conventionally known mast cells mediators released when activated by an IgE-dependent mechanism in allergic rhinitis, mast cells release a variety of cytokines such as IL-4, IL-5, IL-6, IL-8, IL-10, IL-13 and TNF-α [12, 24, 25]. Furthermore, mast cells in patients with allergic rhinitis express higher levels of very-late activation antigen-4 and 5 and mast cell - extra cellular matrix interactions upregulate cytokine secretion from these cells [14]. Such a mechanism may contribute to the enhancement of mast cell activation especially when the levels of antigen in the microenvironment are rather low and contribute to nasal hyperresponsivess [14]. Mast cells also induce IgE synthesis in B cells [28] and as will be described later in this article, mast cells play a key role via the IgE-IgE receptor mast cell cascade [19, 27]. Thus, mast cells are not only effector cells of the immediate phase response, but also act as immunoregulatory cells of the late phase allergic reaction as well as on-going allergic inflammation.
Like mast cells, basophils are also key cells in allergic inf lammation and are derived from pluripotent CD34+ hematopoietic progenitor cells [28]. While human basophils ordinarily do not reside in peripheral tissues and are therefore not detected in the normal nasal mucosa, they are detected in the nasal secretion of patients with allergic rhinitis. The number of basophils in the nasal secretion correlate with the severity of disease. Basophils release histamine, but not tryptase or PGD2, and cytokines like IL-4 and IL-13 in response to FcεRI-dependent activation and are mainly implicated in the late phase allergic response.
Eosinophils play an important role in chronic allergic diseases [29], making them major targets for basic and therapeutic research. Eosinophils derive from the bone marrow from a progenitor cell (CD34+) that may develop into either eosinophils or basophils [30]. Eotaxin appears to be critical for the maturation and release of eosinophils from the bone marrow. Chemokines such as RANTES and eotaxin [30-35] also induce eosinophil recruitment and possibly their activation. Within the tissue, cytokines like IL-5 and GM-SCF keep eosinophils alive for several days or even weeks which overcome programmed cell death (apoptosis) [36, 37]. Mature eosinophils contain MBP [38], ECP [39], eosinophil-derived neurotoxin (EDN) [40], eosinophil peroxidase all of which play important roles in the immune mechanisms of allergic inflammation, especially in altering the surface epithelium. In addition, eosinophils synthesize and release cytokines such as IL-3, IL-5, GM-CSF [41], proinflammatory cytokines, chemokines (RANTES, IL-8, MIP-1α) [42] that play crucial roles in the late phase and on-going allergic inflammation.
Wang et al. [43] studied ECP in nasal secretions in 18 atopic patients and 10 healthy volunteers. Allergen challenge in these atopics induced an increase in eosinophils that persisted for 10 h and was less at 24 h, whereas levels of ECP in these atopics peaked at 24 h indicating possible degranulation [43]. In another study by the same researchers, examined the correlation between the symptoms and the levels of the inflammatory cells in nasal secretions of 18 seasonal allergic rhinitis patients (group 1) outside the pollen season and 40 symptomatic patients (group 2) with seasonal allergic rhinitis during the pollen season. Ten non-allergic subjects (group 3) were also studied as a normal control group. In group 1, 94% patients had an immediate response of nasal symptoms accompanied by a significant increase of histamine, leukotriene C4 (LTC4), and tryptase 5 min after nasal allergen challenge (NAC). One h later, a simultaneous increase was seen both in the percentage of the eosinophils and in the ECP concentration. The eosinophil count reached a peak 2 h after NAC with a duration of 8 h, while the highest ECP level reached only after 24 h with no clear-cut plateau. In group 2, a high percentage of eosinophils was observed. Mostly one observed significantly (p < 0.01) higher concentrations of ECP, LTC4 and histamine but not of tryptase than the baseline values of group 1.
In our study of nasal secretions from 20 patients with allergic rhinitis and 15 patients with infectious rhinitis by transmission electron microscopy we found that the cell population of the allergic group consists of (in order of predominance): eosinophils, neutrophils, lymphocytes, basophilic cells (basophil leukocytes and mast cells), epithelial cells, and macrophages [42]. In the infectious group the population contains: neutrophils, epithelial cells, macrophages, and lymphocytes. Marked degranulation was found in the eosinophils in allergic nasal secretions. Arzuaga et al. [44] studied kinetics of eosinophils in nasal secretion after antigen challenge. They demonstrated that eosinophils of 4.9% at baseline had increased to 13% at the 30 min, 19% at 2 h and 28%, 8 h after the antigenic initiation. On the other hand, the percentage of polymorphonuclear cells was of 40.6%, 34.2%, 34.8% and 31.7% respectively [44].
T lymphocytes are among the principal factors that regulate and co-ordinate immune responses in allergic diseases. Th1 T cells release predominantly IFN-γ and IL-2 and are involved in the delayed hypersensitivity immune reactions, and Th2 T cells release predominantly IL-4 and IL-5 and are predominantly involved in IgE-mediated allergic inflammation. The ligand for the chemokine TARC, CCR4, was found to be expressed by Th2 cells. The attraction of Th2 cells by TARC could represent a key mechanism in allergic reactions as a strong correlation between the levels of TARC in the nasal mucosa and CD4+ CCR4+ cells has been demonstrated [45]. Mucosal inflammation in allergic rhinitis is characterized by the tissue infiltration of T lymphocytes (CD4+ T cells, and CD25+ (activated) T cells) both in the submucosa and the epithelium [46, 47]. There is a significant correlation between the increase in CD4+ T cells during the late phase allergic reaction following an allergen challenge and the number of infiltrating eosinophils in the mucosa [48]. This is associated with an increased expression of IL-3, IL-4, IL-5, GM-CSF at mRNA levels in the nasal mucosa [49]. In perennial rhinitis, there is an increase in CD4+ T memory cells, CD4+ T cells and B cells in the nasal mucosa [47]. This is associated with an increase in the number of IL-4, IL-5 and IL-13 positive cells suggesting a Th2 pattern [50]. Moreover, there is an increase in intra-epithelial γ/δT cells in perennial allergic patients [51]. γ/δT cells are of importance since they are able to regulate allergic immune responses through their capacity to induce IgE synthesis by B cells [52].
The Regulatory T (Treg) cell is one of subsets of CD4+ cells, which suppresses the functions of other lymphocytes and are characterized by surface expression of CD4 and CD25 and nuclear expression of the transcription factor forkhead box P3 (FOXP3). Recently, many functions of forkhead box P3 and its influence on the immune system have been elucidated. T-regulatory cells can influence the development and expression of atopy and the allergic response. Adaptive FOXP3+ Treg cells were essential for establishing mucosal tolerance and for suppressing IL-4 production. Studies have reported about decreased expression of FOXP3 in asthma and allergic rhinitis [53, 54]. The number of Tregs increase after immunotherapy suggesting clearly that Tregs play an important role in inducing tolerance. Clinical improvement seen after allergen immunotherapy for allergic diseases such as rhinitis and asthma is associated with the induction of IL-10 & TGF-β producing Tr-1 cells as well as FOXP3 expressing IL-10 T cells, with resulting suppression of the Th2 cytokine milieu. Activation and expansion of antigen-specific CD4+ CD25+ T(regs) using adjuvants or pharmacological agents such as low dose steroids or vitamin D3 could induce antigen-specific tolerance in allergic diseases. In the nasal mucosa of patients with perennial allergic rhinitis, B cells comprise about 20% of the total lymphocyte population [47] and recent studies have shown that B cells in seasonal allergic rhinitics can undergo class switch to IgE locally in the nasal mucosa.
Allergic reactions occur in a mucosal environment that is rich in both dendritic cells and macrophages. However, there are significant differences between the lower and upper airways, as alveolar macrophages form more than 90% of the cell population in bronchial alveolar lavage [55], but airway macrophages on the nasal epithelial surface just account for about 1 to 2% of the cells. Still, in seasonal and perennial allergic rhinitis, a significant increase in macrophages has also been found in the nose [56]. Langerhans cells represent an important group of dendritic cells in the nose, characterised by the expression of CD1 and Birbeck granules [57]. These cells increase after allergen challenge or in patients with allergic rhinitis [58] and may serve as antigen presenting cells in the upper airway.
Structural cells in allergic inflammation
For many years epithelial cells were considered to play a simple role of a barrier while being involved in the secretion of mucus or removal of foreign agents by their cilia. However, recent studies have shown that epithelial cells have a much wider range of immunomodulatory activities through the release of eicosanoids, endopeptidases [59, 60], cytokines and chemokines (IL-6, IL-8, GM-CSF, TNF-α, RANTES, TARC, eotaxin, SCF) [61, 62] and expression of adhesion molecules like intercellular adhesion molecule-1 and VCAM-1 thus contributing to the enhancement of allergic inflammation. Furthermore, nasal epithelial cells in allergic rhinitics release express and release matrix metalloproteases matrix metalloproteinase (MMP)-2, MMP-9 and MMP-13 [63]. Nasal epithelial cells express HLA-DR, and CD86 and can present antigen to T cells [64]. Also these cells express the alpha and gamma subunits of the FcεRI [65].
It is now appreciated that allergens, on account of their enzymatic proteolytic activity can directly activate cells [66]. House dust mite allergens have been shown to activate epithelial cells in vitro, inducing cytokine and chemokine release and thus can induce airway inflammation independent of IgE [67, 68]. Moroever, Der p1 is able to alter the epithelial tight junctions [69] thereby increasing epithelial permeability. The relative importance of non-IgE to IgE-mediated mechanisms is undetermined. It has also been shown that epithelial cells in allergic individuals are more sensitive to air pollutants like diesel exhaust particles and this has been attributed to the greater constitutive and pollutant induced release of pro-inflammatory cytokines [70].
Epithelial cell-derived thymic stromal lymphopoietin (TSLP) is a master switch for asthma or atopic dermatitis by inducing a dendritic cell-mediated Th2-type allergic inflammation [71]. More recently, studies have shown an overexpresson of TSLP in allergic rhinitis and asthma as well as nasal polyps especially in the epithelium [72-74]. The epithelial TSLP expression was reduced in Fc receptor gamma chain (FcγR)-deficient mice, where the high-affinity IgE receptor (FcεRI) is not expressed on mast cells, in comparison with control C57BL/6 mice indicating that mast cells regulate the epithelial TSLP expression in allergic rhinitis possibly via FcεRI, expression. IL-25 is a member of the IL-17 family. Upregulate IL-25 can amplify on-going allergic inflammation buy upregulating Th2 type inflammation [75]. IL-33 of the IL-1 family also can amplify Th2 type inflammation. Increased levels of IL-33 have been reported in asthma [76].
Infiltration of effector cells is crucial to the development of allergic diseases like asthma and allergic rhinitis. Structural cells like endothelial cells appear to play a dual role in the pathogenesis of allergic rhinitis by participating in the recruitment of leukocytes to the site of the allergic response. Endothelial cell VCAM-1 is overexpressed during the pollen season [77]. Endothelial cells in allergic rhinitics and are also an important source of several cytokines and chemokines like RANTES and eotaxin [78]. Moreover, like epithelial cells, endothelial cells also express the H1 receptor and stimulation with histamine induces activation of these cells [79, 80].
Ongoing inflammation in allergic rhinitis
Structural cells like epithelial cells, residential cells like mast cells and the infiltrated inflammatory cells like eosinophils, basophils and T cells all play a role in inducing and maintaining on-going allergic inflammation. While cytokines like IL-4, and IL-13 released from mast cells and T cells help drive B cells toward IgE synthesis and could contribute to the local IgE synthesis in the nasal mucosa of patients with allergic rhinitis [19, 26, 27]. Most recent studies also have shown that under allergic inflammatory conditions, "primed" mast cells express high levels of the high affinity receptor for IgE and the ligand for the surface antigen CD40, involved in T/B cell interactions leading to IgE production [26, 27]. Mast cells also have the potential to function as antigen presenting cells [81] and IL-4 from mast cells can direct uncommitted helper T lymphocytes toward Th2 and also upregulate the FcεRI expression in mast cells and basophils [26]. These findings suggest that mast cells can orchestrate on-going allergic inflammation. Furthermore, the locally synthesized IgE itself can upregulate the FcεRI expression in mast cells [26, 27]. The augmented FcεRI in mast cells can bind increased number of IgE-Ag complexes which in turn can enhance the sensitivity of mast cells to allergen resulting in the enhancement of the production of immunomodulatory cytokines and chemical mediators, forming an important positive-feedback amplification loop involving the IgE-IgE receptor mast cell cascade (Fig. 2).
Local IgE production
The production of IgE locally in the nasal mucosa is now well established. Studies have shown both by in-vivo allergen change experiments done out of season in ragweed sensitive patients that there is IgE class switch and increase in IL-4 positive cells [82]. Similarly in-vitro allergen challenge studies have shown the upregulation of IgE producing B cells [83]. The locally produced IgE then can play a key role in mast cell activity amplification via upregulating the FcεRI expression in mast cells and subsequent mediator release after crosslinking of the bound IgE [27]. Recent studies have also shown the presence of local IgE in idiopathic rhinitis suggesting the possibility of local IgE production playing a more important role in on-going allergic inflammation [84].
Nasal hyperresponsiveness
Patients with allergic rhinitis report increased symptomatic responsiveness to environmental irritants such as tobacco smoke compared with nonatopic individuals. A hallmark of allergic rhinitis is hyperresponsiveness of symptomatic individuals to specific allergens, a phenomenon known as 'priming', and to nonspecific stimuli such as irritant particles and strong odors; however, the causes of this effect remain unclear. Activated inflammatory cells may foster mucosal penetration by allergen and provide additional targets for antigen-specific stimulation. Increased generation of inflammatory mediators also are expected to do so. In symptomatic patients, hyperresponsiveness to nonspecific irritants may reflect interactions among inflammatory cellular influx, epithelial injury, and increased end-organ responsiveness caused by exposure to an antigen.
Nerve growth factor (NGF) which is over expressed in allergic rhinitis is linked to the development of hyperresponsiveness, and is also associated with allergic inflammation, thus bridging the gap between these two features of allergic rhinitis and asthma. NGF is synthesized, stored, and released by cells that play key roles in the allergic inflammatory disease of the airways, including mast cells, eosinophils, CD4+ T cells, B cells, and respiratory epithelial cells [85]. Mast cells express and secrete NGF [85, 86], and they have been shown to be in close apposition with tachykinin-containing nerve endings in peripheral tissue [87]. Other cells in the later phases of inflammation may also contribute to NGF release in the human airways, based on NGF elevation in bronchoalveolar lavage fluids 18 h after segmental allergen challenge in asthmatics [88].
Minimal persistent inflammation
There is evidence of ongoing inflammation in the airway even when rhinitis or asthma symptoms are quiescent. This has had important implications for the way we treat and has led to the concept of the need for long term anti-inflammatory treatment for those with rhinitis and asthma [89].
Systemic inflammation in allergic rhinitis
AR is not associated with just localized inflammation of the nasal mucosa but also has a systemic element to it. This may explain in a significant way the mechanisms underlying the rhinitis-asthma link. In individuals with seasonal allergic rhinitis without co-existent asthma, nasal allergen provocation produced increased adhesion molecule expression, eosinophil infil tration in both the upper and lower airways, and increased bronchial hyper-reactivity [90, 91]. These results demonstrate that an allergic nasal reaction produces systemic inflammatory changes. In sensitized subjects, allergen exposure activates immune cells, including Th2 lymphocytes, dendritic cells, mononuclear cells, mast cells, and others, both within the nose and in nasal-associated lymphatic tissues. Some of these Th2 cells migrate to the bone marrow, where they stimulate the bone marrow to produce and recruit the inflammatory cells, including basophils, eosinophils and mast cell precursors, to the inflamed target tissues.
Link between allergic rhinitis and asthma
The increasing evidence on the links between AR and asthma comes from epidemiologic, immunologic, and clinical studies. Epidemiologically, up to 40% of patients with AR also have asthma, and up to 80% of patients with asthma experience nasal symptoms. Furthermore, patients with AR are at three times the risk of developing asthma compared with those without AR and in children who develop rhinitis within the first year of life the chances of developing asthma are twofold greater as compared with those who develop rhinitis later in life. Therapeutic outcomes of treating AR in asthma have shown that AR treatment improves asthma symptoms and lowers overall costs and reduces hospitalizations [92]. In a recent international survey on the impact of concomitant AR and asthma on patient health and quality of life, the presence of AR in children with asthma disrupted their lives by limiting their ability to get a good night's sleep (79% of adults and children), participate in leisure and sports (75% of adults and children), concentrate at work/school (69% of adults and 73% of children), or enjoy social activities (57% of adults and 51% of children), worsened [93].
Both AR and asthma are inflammatory diseases and their inflammatory mechanisms are similar in that they are characterized by an inflammatory infiltrate made up of eosinophils, T cells, and mast cells that release several mediators, chemokines and cytokines, local and systemic IgE synthesis, and a systemic link via the bone marrow. Typical early- and late-phase responses are also cardinal features common to both rhinitis and asthma. Studies have shown that patients with AR exhibit bronchial hyperresponsiveness and increase in inflammatory cells, and that nasal allergen challenge further increases this hyper-reactivity. Eosinophils have been demonstrated in the nasal mucosa of patients with asthma, even in the absence of symptoms of rhinitis. In addition, patients with AR have increased number of inflammatory cells in their bronchial mucosa. This increase in bronchial inflammation in response to allergen-induced rhinitis might contribute to the exacerbations of asthma frequently seen in individuals with underlying AR. Allergen challenge increased the bone marrow concentrations of both cytokines and progenitor cells both in animal models and in individuals with atopic asthma and in addition, in atopic individuals who had late asthmatic responses and airway eosinophilia, allergen inhalation caused trafficking of T lymphocytes to the bone marrow, enhancing the ability of these lymphocytes to generate IL-5. Ultimately, these newly generated inflammatory cells enter the circulatory system, from where they are selectively recruited to the target organs (lungs and nose), exacerbating airway inflammation. In addition to stimulating inflammatory cell production, cytokines upregulate the expression of adhesion molecules, further facilitating the recruitment of inflammatory cells into nasal and bronchial tissues. This selective recruitment of inflammatory cells into the airways would occur only in individuals with pre-existing asthma in whom specific adhesion molecules, such as VCAM-1, and chemoattractants, such as eotaxin, already exist. Persons without asthma do not have these addressins in their airways and thus do not have the machinery in place to recruit inflammatory cells into their airways during exacerbations of rhinitis. This parallel relationship is influenced by many interactions between the nose and the lower airways: some interactions stem from the fact that the nasal passages play a major homeostatic role by conditioning inhaled air, but perhaps even more important is the bidirectional interaction that results from the systemic inflammation that is produced after local allergic reactions.
CONCLUSION
Allergic rhinitis is characterized by an inflammatory infiltrate made of different cells. This cellular response includes the chemotaxis, selective recruitment and transendothelial migration of cells, the localization of cells within the different compartments of the nasal mucosa, the activation and differentiation of various cell types as well as a prolongation of their survival, the release of mediators by these activated cells, the regulation of the local and systemic IgE-synthesis and the communication with the immune system and the bone marrow. These events take place only in subjects who have been sensitized to allergens before, i.e. allergen-specific IgE-antibodies have been formed and bound to the membrane of mast cells and other cells, but not in healthy individuals, who do not show a measurable reaction of the nasal mucosa to the same allergens.
Although, histamine, (released by mast cells and basophils), is still one of the major effectors of the allergic reaction, with the increasing knowledge and better understanding of the mechanisms of the allergic reaction, many other mediators produced by different cell types are found to be involved. Thus, the intricate interaction amongst these mediators, cytokines, chemokines, neuropeptides, adhesion molecules and various cells in the form of a complex network lead to the onset of specific symptoms and the non specific hyperreactivity of allergic rhinitis. The pathophysilogy of allergic rhinitis may be further complicated by the involvement of neural reflex mechanisms and interaction of allergic and non-allergic stimuli.
The vast majority of patients with asthma have rhinitis, and rhinitis is a major independent risk factor for asthma in cross-sectional and longitudinal studies. The relationships between rhinitis and asthma can be viewed under the concept that the two conditions may be manifestations of one syndrome, the 'chronic allergic respiratory syndrome', occurring in two different parts of the respiratory tract. Some interactions arise from the fact that the nasal passages play a major homeostatic role by conditioning inhaled air, but perhaps even more important is the bidirectional interaction that results from the systemic inflammation that is produced after local allergic reactions. Although there are cardinal structural differences between the nose and the lungs, these experimental and real-life observations on the link between rhinitis and asthma outweigh the differences and lend support to the concept that rhinitis and asthma may be considered as manifestations of one syndrome that has a wide spectrum of severity. Successful management of this chronic allergic respiratory syndrome requires an integrated view of the airways, understanding of their interactions, and an integrated approach of treatment also targeting systemic inflammation.
Finally, the understanding that the mechanisms of disease generation provide a framework for rational therapy in this disorder, which will be based on the complex inflammatory reaction rather than on the symptoms alone is very important to the development of newer and more effective strategies of treatment.

 XML Download
XML Download