

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Lipocalin-2 (LCN2) is a 25 kDa secreted glycoprotein belonging to the lipocalin family which contributes to the transport of small hydrophobic molecules [12]. LCN2 participates in the control of various biological processes such as cell death [3], cell migration [4], cell differentiation [5], and innate immunity [67]. It is also known as neutrophil gelatinase-associated lipocalin (NGAL), because it was first identified from human neutrophils [8]. LCN2 is expressed in multiple tissues, i.e., kidney, liver, adipocytes, bone marrow, and uterus [910]. LCN2 was suggested as a sensitive biomarker for various forms of kidney injuries as LCN2 in the renal tubules is highly expressed about 1,000-fold, and released into plasma under infection, or ischemic condition [11121314]. Several studies also reported that circulating levels of LCN2 are associated with obesity, metabolic syndrome, diabetes, cardiovascular disease, and cancer [1516]. Recently, the receptors of LCN2 were found in microglia [17], astrocytes [18], neurons [19], and choroid plexus [20]. LCN2 secreted by glial cells regulates neuronal morphological changes [17], contributes to neuronal cell death [21], is associated with reactive astrocytosis [18] and neuroinflammation [22], and act as a chemokine inducer in the central nervous system (CNS) [23]. Among the CNS disease, the risks of experimental autoimmune encephalomyelitis [24], intracerebral hemorrhage [25], and spinal cord injury [26] was reported to be positively associated with circulating LCN2 levels. However, the role of LCN2 in the CNS has not been fully elucidated yet.
In this paper, we particularly reviewed the significant evidences showing that LCN2 is implicated in Alzheimer's disease (AD) [2728] which may suggest that LCN2 could control neuroinflammation [29], insulin signaling [30], neurite maturation [31] related to AD pathogenesis. This review may provide the possibility of LCN2 for the regulation of the AD pathogenesis.
THE ROLE OF LCN2 IN THE BRAIN
LCN2 known as NGAL [32], or growth factor-stimulated inducible protein 24 [33] is a 25 kDa glycoprotein purified from human neutrophils [34]. LCN2 is composed of extracellular ligand-binding proteins with a high specificity for hydrophobic molecules [3536]. LCN2 binds to 2 specific receptors such as brain type organic cation transporter (24p3R) and megalin [143738]. The 24p3R exists not only in the neutrophils [39] particularly with high levels, but also in the microglia [17], astrocytes [18], and neurons [19]. Megalin is mainly expressed in the brain capillaries, choroid plexus [20], the ependymal cells of the lateral ventricles [40], neural tube [41], astrocytes [42], and neurons [4344]. Previous studies demonstrated that lipocalins acts as a critical role in multiple physiological processes including immune responses, cell migration, proliferation [45], and the regulation of cell homeostasis against exogenous compounds [2]. Particularly, LCN2 promotes neuronal death [46] and reactive gliosis [224748], and triggers insulin resistance (IR) [30] in the CNS. Recently, several studies suggested positive relationship between LCN2 and pathophysiology of AD [2849]. Increased LCN2 levels were observed in the postmortem of AD brains [28] as well as in people with mild cognitive impairment (MCI) [50]. Based on these evidences, alteration of LCN receptors and the increased level of LCN2 in the CNS may contribute to the progression of AD which may be related to neuronal cell death, glia's activation [51], and IR [52]. Even though the role of LCN2 in AD brain was not fully understood until now, recent studies suggest that LCN2 would be a potential marker for AD progression.
LCN2 CONTRIBUTES TO NEUROINFLAMMATION IN THE AD BRAIN
Neuroinflammation aggravates the pathogenesis of AD
Neuroinflammation is known to be mediated by microglia as the resident brain macrophage, astrocytes, neurons, neutrophils, and various inflammatory mediators released from these cells [51]. In the initiation state, neuroinflammation is beneficial for repairing the damaged tissue, but the excessive inflammatory responses are detrimental for neuronal regeneration [53]. Recent studies demonstrated that excessive neuroinflammatory response could impair the neuronal circuit in the CNS [54], and chronic neuroinflammation is critical for the onset and the progression of neurodegenerative diseases including AD [5556]. Moreover, current reports indicate that high levels of inflammatory mediators have influence on diverse regions in the brain, and contribute to the cognitive dysfunction in the AD [575859]. The study on neuroinflammatory mechanisms in AD brain is needed to ascertain the strategies to prevent the onset and the progression of AD.
LCN2 contributes to the inflammation in the brain
It has been reported that LCN2 contributes to the immune responses leading to the pathogenesis of neurodegenerative diseases [28294760]. Neuroinflammation by the brain injury [61] or infection [62] could promote the secretion of LCN2 from astrocyte, microglia, endothelial cell as well as neurons [25]. Released inflammatory mediators from the activated microglia and astrocytes play a critical role in cell migration and recruitment of these glial cells to the injury site [6364]. LCN2 accelerates neuronal motility and its morphological alteration [21], and cell migration in glial cells and endothelial cells [2365]. LCN2 promotes phenotypic changes of glial cells by a Rho-ROCK (Rho kinase) [31], and the migration of astrocyte, microglia and neurons by secreting chemokines such as CXCL10 [65]. Moreover, chronic and excessive immune response triggers the over-secretion of inflammatory cytokines including LCN2 in the CNS, subsequently aggravates the neural imbalance in hippocampus, and leads to the impaired long-term behavior [666768]. In fact, increased LCN2 levels were found in the brain of the AD [286169], and the MCI [50]. Considering the previous findings, LCN2 contributes to the neuroinflammation in the brain, and is especially linked to the neuroinflammatory responses in the AD brain. Given that the neuroinflammation contributes to the aggravation of AD pathogenesis [5457], further studies are needed to elucidate the role of LCN2 on inflammatory mechanisms in AD brain.
LCN2 CONTRIBUTES TO IR IN THE AD BRAIN
IR aggravates the pathogenesis of the AD brain
Several epidemiologic studies have shown that type 2 diabetes [70], and demonstrated that IR as impaired insulin mechanism is related to the pathogenesis of cognitive declines in the AD [717273]. Previous studies suggested that impaired insulin signaling was found in the postmortem of AD brains [5274]. Normally, insulin signaling contributes to the inhibition of beta amyloid (Aβ) oligomerization and tau phosphorylation in the brain [52] as well as the transportation of Aβ out of the brain [75], while impaired insulin signaling leads to the pathogenesis of AD [7677]. One in vitro study demonstrated that reduced activations of the insulin receptor/insulin receptor substrate 1 (IR/IRS1)/PI3K/AKT, and insulin growth factor-1 receptor (IGF-1R)/IRS2/PI3K signaling were found in the AD brains [74]. Moreover, disrupted insulin signaling is negatively associated with the volumes of hippocampus [71], medial prefrontal cortex, and medial temporal area [78]. Additionally, IR results in the disconnection in the cerebellum-frontal-temporal cortex [79]. Recent studies demonstrated that the site of IRS1 phosphorylation which is important to identify IR [8081], is a key in the progression of AD [82]. Recent studies suggested that IR may be one of important causes which lead to diabetes induced dementia involving to cognitive decline [910]. Collectively, IR is a critical issue in the AD, and should be highlighted to find the mechanisms related to the onset and progression of AD.
LCN2 is associated with IR in the brain
Considering the previous evidences, circulating LCN2 levels were associated with hyperglycemia, IR, metabolic syndrome, diabetes, cardiovascular disease [1630838485], in vitro [8687], in vivo [8388], and clinical researches [89] reported that LCN2 is involved in the negative regulation of insulin sensitivity, and could directly reduce the insulin sensitivity. Knockdown of LCN2 gene in the adipose tissue was reported to reduce IR closely related to obesity and diabetes [30]. Increased levels of LCN2 in both circulation and adipose tissue were observed in the obese and type 2 diabetic patients [1683]. These evidences suggest that the reduction of LCN2 levels may contribute to the improvement of IR in the AD although the relationship between LCN2 and IR in the brain was not fully elucidated. Taken together, specific mechanisms on the association between LCN2 and IR in AD brain should be further investigated, which may suggest that LCN2 could aggravate IR which progress AD [3086].
LCN2 CONTRIBUTES TO THE COGNITIVE DECLINE
Recent studies demonstrated that LCN2 is involved in some behaviors such as cognitive functions, emotional behaviors, depression, and anxiety [689091]. Especially, LCN2 plays a role as a pivotal regulator in the cognitive behaviors [90]. As the LCN2 gene expression was modulated by glucocorticoids which is related to cognitive function [92], and high levels of LCN2 were found in patients with MCI dementia [4750], high levels of LCN2 may lead to the cognitive decline. It was reported that high levels of LCN2 triggers inflammation, and subsequently reduces cognitive function [4850]. Moreover, LCN2 could regulate the formation of dendritic spines and maturation of dendrites [93]. It could also control the neuronal morphology and plasticity by transporting iron which suggested that synaptic plasticity is related to iron metabolism [94]. Based on the significant evidences, LCN2 is an important factor to regulate cognitive function in the AD. Furthermore, the association between LCN2 and IR in the brain also supports the possibility of LCN2 to control the cognitive decline [69].
CONCLUSION
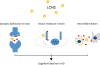
In this review, we highlight 3 points: first, LCN2 could aggravate inflammation in the AD brain; second, LCN2 is involved in IR in the AD brain; and third, LCN2 could contribute to cognitive decline in the AD by modulating inflammation and controlling synaptic plasticity (Figure 1). Taken together, we suggest that the modulation of LCN2 may be a key to attenuate the pathogenesis of AD. In addition, further studies such as in vivo study using transgenic animal model, and long-term clinical studies are needed to decipher the role of LCN2 in the pathogenesis of AD and cognitive impairment.
Figure 1
The schematic image regarding the role of LCN2 in AD brain. LCN2 is involved in the neuroinflammatory responses in AD brain, and may aggravate insulin resistance in the AD brain, and cognitive decline in the AD by controlling inflammation and synaptic plasticity.
LCN2, lipocalin-2; AD, Alzheimer's disease.
 XML Download
XML Download