

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Lancefield group B streptococci (GBS), also referred to as Streptococcus agalactiae, is a gram-positive, opportunistic pathogen that colonizes the gastrointestinal and genitourinary tracts of up to 50% of healthy adults [123]. In 1938, it was first identified as a human pathogen, causing human fatal puerperal sepsis [4], but remained relatively unknown as sporadic asymptomatic cases were reported until the 1960s. By the 1970s, GBS had emerged as the predominant pathogen causing septicemia and meningitis in neonates and infants living in diverse regions [5678910]. GBS infection in newborns is usually classified as an early-onset disease (EOD) and late-onset disease (LOD), respectively) depending on the age of the infant at the time of disease manifestation [11]. Recent advances in the diagnosis and treatment of GBS infections and global hygiene standards have significantly reduced the development of neonatal infections and mortality, particularly due to EOD. However, recent estimates also show 0.5–2 cases of neonatal GBS infections per 1,000 births with a mortality rate of 9.6%–22% [1213]. In addition, recent reports have revealed that an increasing number of those infections occurred in pregnant women and non-pregnant adults who typically had an underlying medical condition. The incidence of GBS infection among those adults increased from 3.6 cases/100,000 persons in 1990 to 7.3 cases/100,000 persons in 2007, with significantly higher case fatality rate at 15% [14]. Although vaccination is the most promising strategy for preventing GBS infection, currently no licensed GBS vaccine is available in the market. Thus, the development of a GBS vaccine is the need of the hour, considering the risk involved in presently used prenatal strategies and the prevalence of GBS infections in the elderly.
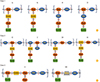
GBS are covered with a sialic acid-rich capsular polysaccharide (CPS) and belong to one of the ten serotypes (Ia, Ib, and II–IX). Each CPS consists of variously arranged monosaccharides and a sialic acid residue on the branching terminus of the repeating unit (Fig. 1) [1516]. Similar to that of pneumococci, GBS CPSs also show potential immune evasion mechanism for GBS by inhibiting complement deposition and phagocytosis [1718]. Recent systemic meta-analyses indicate that five serotypes (Ia, Ib, II, III, and V) account for 97% of invasive isolates in all geographical regions [19]. Owing to its importance in GBS pathogenesis, CPS is considered to be the prime vaccine candidate for GBS.
In 2014, World Health Organization convened the first meeting of the Product Development for Vaccines Advisory Committee for consultation regarding the development of GBS vaccines [20]. In this meeting, they agreed that native CPS vaccine is ineffective due to its poor immunogenicity, but the immunogenicity of the GBS polysaccharide conjugate vaccine (PCV) may be able to induce a stronger and higher functional CPS-specific IgG response [2122]. Novartis has developed a trivalent (Ia, Ib, and III) CRM197 conjugate vaccine, and conducted a phase 1b/2 clinical trial (NCT01193920) in infants born to women who were vaccinated with a trivalent GBS PCV [23]. In infants born to the GBS PCV recipients, the level of CPS-specific antibody was higher at birth than at 43 and 91 days, indicating that maternal GBS PCV immunization is intended to protect infants by sufficient CPS-specific antibody transfer across the placenta. In 2017, Pfizer started to evaluate a pentavalent GBS PCV targeting Ia, Ib, II, III, and V in a phase 1 trial on healthy volunteer (NCT03170609). Despite the promising result from the clinical trial, recent change of serotype distribution worldwide requires the replacement of old serotypes, or the addition of new serotypes in the GBS PCV.
Although multivalent CPS PCVs are well established in terms of their production, safety, and immunogenicity, it has several limitations and various concerns have been raised regarding its use. The limitations are immune interference with similar type of conjugate vaccines, including Haemophilus influenzae type b, meningococcal, and pneumococcal conjugate vaccines, potential problems of serotype replacement and switching, and diverse serotype distribution across and within geographical regions [24252627]. In addition, an increasing number of reports show that nonencapsulated GBS strains are a concern for the implementation of an anti-CPS vaccine [28293031]. Structurally conserved protein antigens that are essential for GBS virulence and can induce a strong immune response against most of the GBS strains, are emerging as the most attractive and cost-effective vaccine candidates [12323334]. MinervaX Inc. recently announced that their protein-only vaccine based on the fusion of highly immunogenic N-terminal domains of Alpha C and Rib (GBS-NN) showed positive results from a phase I trial in 240 healthy adult women [35]. All subjects immunized with one or two doses of GBS-NN showed an increase of over 30-fold in GBS-NN specific antibodies compared to pre-immune level [36]. GlaxoSmithKline also identified a conserved sequence encoding components of GBS pili proteins, which induced the immune response against different GBS serotypes in preclinical studies [3233]. We investigated that the C-terminal end of a serine-rich repeat surface glycoprotein named latch domain could provide serotype-independent protection in mouse meningitis model [37]. In addition, many surface proteins of GBS are being investigated, at the pre-clinical stage, as broad spectrum vaccines [3538394041424344454647].
Because of the possibly low baseline incidence of the primary endpoint of invasive disease, there is an urgent need for a standardized clinical efficacy assay for GBS vaccines in order to support and accelerate the clinical studies. In pneumococcal PCV, two standard immunological methods, enzyme-linked immunosorbent assay (ELISA) and multiplex-opsonophagocytosis assay (MOPA) for measuring CPS-specific antibody and functional antibody titers, are well established and accepted as standard vaccine efficacy assays. The radio-antigen binding assay (RABA) has been the gold standard for the quantification of anti-GBS antibody as it measures antibody in its native state [48]. However, the RABA has several limitations, such as low detection sensitivity, limited ability to quantify Ig isotypes, and the difficulties of procuring and using radioisotopes. Several ELISA protocols that are more sensitive and isotype-specific have subsequently been developed based on pneumococcal CPS ELISA. However, the sensitivity and non-specific binding remain a concern for these methods as well [495051]. It is important to note that the results of RABA and ELISA might not always reflect functional antibody level for encapsulated bacteria, as experienced in pneumococcal vaccines [52].
In vitro opsonophagocytic assay (OPA) is believed to have a reliable correlation with the functional efficacy of pneumococcal PCVs, because host protection against pneumococcus is mainly mediated by opsonin-dependent phagocytosis [5354]. As pregnant women are mainly immunized with the GBS vaccine, and as the maternal transfer of anti-polysaccharide (PS) specific IgG should be tested using extremely small amounts of serum from the newborns, our group developed, standardized, and validated three-fold multiplexed GBS-OPA (GBS-MOPA) to enable practical, large-scale assessment of GBS vaccine immunogenicity [55]. Therefore, the standardized functional efficacy assay would be essentially used to evaluate the clinical efficacy in the process of GBS PCV approval and licensure.
Application of a GBS vaccine is the most promising strategy for the prevention of GBS infections in both newborns and adults with underlying diseases. However, numerous questions arise during the designing and evaluation of GBS vaccines. First, recent epidemiology studies have introduced the phenomenon of serotype switching and replacement occurring worldwide [25565758]. Thus, a streamlined effort is needed to update the global disease burden estimate and serotype distribution. Second, a standardized immunological assay is urgently needed. Although we developed and standardized GBS-MOPA, it needs to be further optimized to suit the specific needs of different countries. In addition, standard ELISA to quantify CPS-specific antibody should be characterized by multi-national based assessment. This review will support the development of a new strategy for GBS vaccine development, evaluation to substantially reduce the global burden of GBS infections, achieve substantial reduction in premature and still births worldwide.
 XML Download
XML Download