

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Severe fever and thrombocytopenia syndrome virus (SFTSV) is a Phlebovirus belonging to the Bunyaviridae family [1]. This family is comprised of a group of segmented and negative-strand RNA virus with more than 350 viruses grouped into to five genera: Orthobunyavirus, Hantavirus, Nairovirus, Phlebovirus, and Tospovirus. Severe fever and thrombocytopenia syndrome (SFTS) was one of the newly discovered virus from Phlebovirus [1] which has now over 70 antigenically distinct serotypes wherein 68 of the known serotypes are divided into two groups: Phlebotomus fever viruses, which includes 55 virus members transmitted by Phlebotominae sandflies, and Uukuniemi viruses, which are transmitted by ticks and are comprised of 13 members [23]. Aside from SFTSV, eight species of the genus Phlebovirus, including Alenquer, Candiru, Chagres, Naples, Punta Toro, Rift Valley fever, Sicilian, and Toscana viruses, cause human disease [45]. Further, SFTSV is distantly related to both the existing Uukuniemi virus and Phlebotomus fever viruses, which is why it was classified as a Phlebovirus [126].
SFTSV is a spherical virion of 80–100 nm in diameter covered by a lipid bilayer envelope of 5–7 nm in thickness [17] and is known to contain a tripartite RNA genome which has three single-stranded RNA segments: small (S; 1,744 bp), medium (M; 3,378 bp), and large (L; 6,368 bp) [16]. The L segment encodes the RNA-dependent RNA polymerase, the M segment encodes the viral envelope glycoproteins (Gn and Gc), and the S segment encodes both a nucleoprotein and a nonstructural protein in an ambisense orientation (Fig. 1) [1].
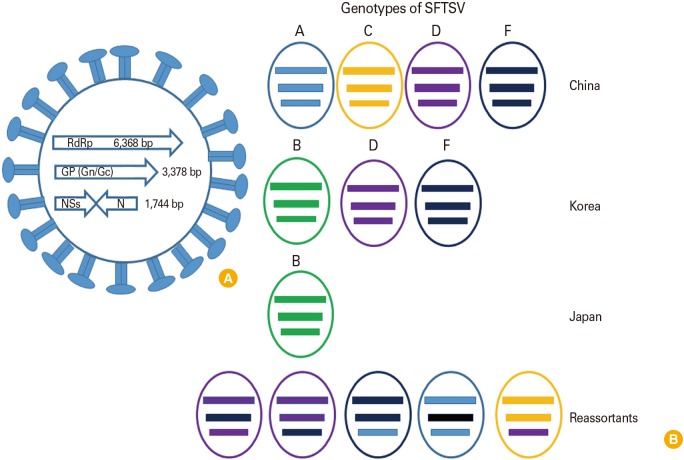 | Fig. 1Genetic composition of severe fever and thrombocytopenia syndrome virus (SFTSV) and genetic diversity. (A) SFTSV has three segmented RNA genomes (L, M, S). L and M segments have one open reading frame with negative sense polarity. However, S segment has ambisense RNA encoding two viral proteins, the nonstructural proteins (NSs) and N proteins. (B) To date, A to F genotypes of SFTSVs were reported. Of these, genotype A, D, and F were the most commonly observed in mainland China. Genotypes B, D, and F were co-circulating in South Korea and genotype B was the most prevalent strain in Japan. Due to the segmented nature of SFTSV, genetic reassortments which occurred resulted in unique genotypes. RdRp, RNA-dependent RNA polymerase.
|
Hospitalized SFTS patients have clinical symptoms that include fever, thrombocytopenia, gastrointestinal symptoms, and leukocytopenia [1]. Laboratory tests of SFTSV cases commonly show elevated serum levels of alanine aminotransferase, aspartate aminotransferase, blood urea nitrogen, lactate dehydrogenase, creatinine kinase MB fraction, and increased activated partial-thromboplastin time [1]. SFTS can be traced back to initial reports of an emerging infectious disease in rural areas of Hubei and Henan provinces in Central China in 2009 [1]. However, at the time of infection the causative agent had not been determined, and due to similar clinical manifestations, the differential diagnosis includes Anaplasma phagocytophilum infection [1], hemorrhagic fever with renal syndrome, and Rickettsia tsutsugamushi infection [8]. Active surveillance and a thorough investigation including laboratory testing, viral isolation, cell line cultures, and molecular characterization were then implemented to properly diagnose the disease resulting in the isolation and confirmation of SFTSV in a 42-year-old farmer from Henan, China [1]. Surveillance data showed that SFTSV spread to at least 10–15 provinces in China in 2010–2013 [4]. Furthermore, SFTS cases have been reported in other Asian countries including South Korea and Japan and as well as in Mediterranean countries and the United States [9101112].
Go to : 
Epidemiology and Vectors of SFTSV
The majority of SFTS cases have been identified in East Asia, primarily in China, Korea, and Japan [1131415]. Out of the 7,419 cases of SFTS reported before 2016, there were 355 estimated total deaths in China, and at this time the virus was mainly distributed in Eastern and Central China [16]. The first reported SFTS case in Japan occurred in Yamaguchi in 2012 [17] with another 11 positive cases identified after a retrospective study was conducted in the Western provinces [14]. By the end of 2013, the total number of SFTS cases was increased to 40 by infectious disease specialists in Japan, though misdiagnosis remains a possible cause of the low number [17]. SFTSV also occurred in South Korea where the first clinically suspected patient was reported on Jeju Island in May 2013, followed by SFTSV isolation by Seoul National University Hospital in a retrospective study [1518]. The Korean Center for Disease Control reported that until November 2013, there were 36 positives out of 404 suspected SFTSV cases, wherein 17 of 36 confirmed cases were fatal cases [19]. Epidemiological studies in these countries revealed that 90% of patients were older than 35 years of age (median age of 58 years) and most were farmers [19]. In addition to reports in Asian countries, SFTS was also reported in the United Arab Emirates in 2011, although the positive patient was of Asian lineage and a North Korean resident. Thus, SFTS infection is also suspected to be present in North Korea [9].
SFTS is classified as a tick-borne disease with Haemaphysalis longicornis as the predominant vector and Rhipicephalus microplus also acting as a vector based on prevalence studies in epidemic and non-epidemic areas [2021]. H. longicornis predominantly feed on farm animals including goats, sheep, cattle, and horses, but can also be found on companion animals including dogs and cats [22]. In human cases, tick bites before the onset of clinical symptoms were reported [1323]. Based on previous studies, there was a high percentage of sequence homology (95%–100%) between SFTSV isolates collected from humans and animals in areas where SFTSV was endemic [2] and the seroprevalence of SFTSV antibodies in mammals like goats, cattle, and dogs was up to 57% [24]. Thus, the tick-born nature of this pathogen makes it a major threat to both humans and animals. Moreover, human to human transmission of SFTSV has been demonstrated and found to occur through close contact with the blood or body secretions of infected patients [2526]. Therefore, people working in rural areas, especially farmers, hospital staff attending to positive SFTSV patients and those with high levels of interaction with animals are at risk of acquiring SFTSV infection.
Go to :
Genetic Diversity of SFTSV
To investigate the genetic relationship of co-circulating SFTSV strains in specific regions, the viral genome was phylogenetically analyzed. Originally, the nomenclature for SFTSV genotypes was assigned based on the country of origin, such as C1 to C5 (Chinese lineage) and J1 to J3 (Japanese lineage). However, based on full-length sequencing of multiple SFTSVs (159 strains) including those from China, Korea, and Japan, Fu et al. [27] have presented new nomenclature for SFTSV which divides the virus into six clades referred to as genotypes A to F. In this review, we have adopted this nomenclature genotypes A to F. Out of a total of 159 SFTSV strains, 149 were classified into the genotypes A, B, D, E, and F. Of these A, D, and F were the most commonly observed in mainland China, while genotype B was commonly found to be circulating in Korea and Japan. Moreover, genotypes B, D, and F were co-circulating in South Korea and genotype B was the most prevalent strain in Japan. Due to the segmented nature of SFTSV, genetic reassortments which occurred in China and Japan resulted in ten strains with unique genotypes [2728]. Of these, four strains had L and S segments from genotype D with the M segment from genotype F (abbreviated as DFD) while the other six strains were a mixture of genomic segments (DDF, FFA, AFA, FAF, DDF, and CCD) (Fig. 1).
Thus, since SFTSV is an emerging infectious zoonotic disease, understanding the geographic distribution, genetic diversity, and prevalence of SFTSV genotypes will require the use of unified nomenclature and collaborative international surveillance of SFTSV.
Go to :
SFTS Virus Incidence in Animals
SFTSV is primarily transmitted by H. longicornis which may feed on animals including rodents, deer, cattle, goats, dogs, and cats [23] thereby increasing the epidemic threat of SFTSV due to the close interaction of these animals and humans. However, the limited clinical signs of SFTSV infection in animals makes it difficult to identify and study the etiology of this virus in animals [29]. A study conducted in Jiangsu and Shandong provinces in China showed that many domestic animals including goats, cattle, chicken, pigs, and other animals can be infected by SFTSV without any, or with only suspected clinical symptoms (Table 1) [28].
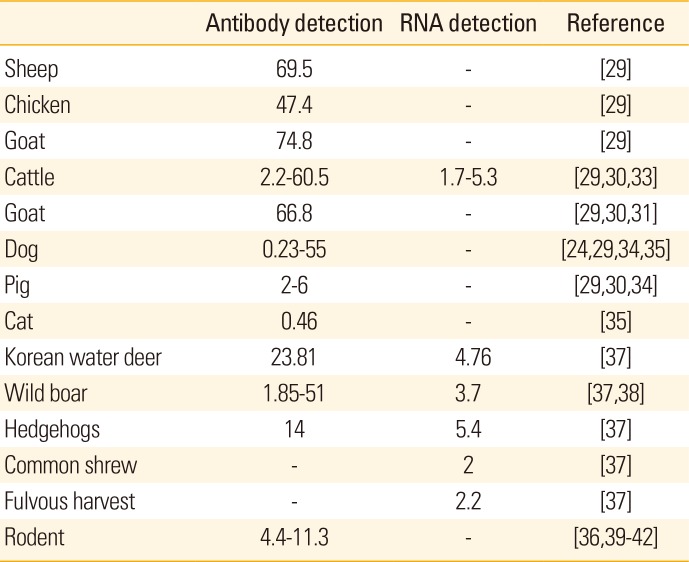
Table 1
Percentage of sero-prevalence and RNA detection in animal models
| Antibody detection | RNA detection | Reference | |
|---|---|---|---|
| Sheep | 69.5 | - | [29] |
| Chicken | 47.4 | - | [29] |
| Goat | 74.8 | - | [29] |
| Cattle | 2.2–60.5 | 1.7–5.3 | [29,30,33] |
| Goat | 66.8 | - | [29,30,31] |
| Dog | 0.23–55 | - | [24,29,34,35] |
| Pig | 2–6 | - | [29,30,34] |
| Cat | 0.46 | - | [35] |
| Korean water deer | 23.81 | 4.76 | [37] |
| Wild boar | 1.85–51 | 3.7 | [37,38] |
| Hedgehogs | 14 | 5.4 | [37] |
| Common shrew | - | 2 | [37] |
| Fulvous harvest | - | 2.2 | [37] |
| Rodent | 4.4–11.3 | - | [36,39,40,41,42] |
![]()
In several studies, majority of patients suffering from SFTS were identified as farmers whose living depends on farm animals [130]. For example, in Yiyuan County in Eastern China, most of the farm families have herds of goats, which are typically infested with hundreds of ticks, of which 83% (111/134) tested positive for SFTSV antibodies [24]. Moreover, in a natural infection study done in Jiangsu Province, China; five goats that were allowed to roam the grasslands were found to become viremic within a short period, and they did all sero-convert without any mortality [31]. Further, SFTSV antibody was found most frequently in goats, with 83/111 (74.8%) being positive out of tested animals in Shandong Province [7]. Next to goats, surveillance reports showed that cattle displayed the highest seroprevalence of SFTS in China with 57.1%–60.5% positive [32]. Cattle are also prone to SFTSV due to their grazing in grasslands which increases exposure to ticks carrying the virus. In Shandong Province, 509 of 842 cattle (60.5%) surveyed were positive for SFTSV-specific antibody; however, the SFTSV RNA detection rate was low as ranged from 1.7% to 5.3% [29]. Phylogenetic analysis of the virus isolates obtained from cattle showed >95% homology with SFTSV isolates obtained from patients in the same area suggesting a possible link between SFTSV infections in humans and domesticated animals [28]. In Japan, serum collected from 510 slaughtered cattle was tested for anti-SFTSV antibodies and 11 samples, mostly from the Shimane Prefecture, were found to be positive [33]. In contrast, pigs displayed only 2%–6% seroprevalence based on a study in various endemic regions in China [34] and in the serosurvey conducted in Shandong Province by Niu et al. [29] only 3.1% (26/839) of pigs were positive for SFTS-specific antibodies.
Companion animals like dogs and cats may also become infected with H. longicornis, which could markedly affect transmission of SFTSV from animals to humans. The SFTSV seropositive rate of dogs was found to vary greatly with seropositivity in Shandong Province of 37.9% (136/359) [29], 6% in Jiangsu [24], and 55% (6/11 dogs) in Hubei, China [34]. In South Korea, a surveillance study of animal shelters identified SFTSV infection in a single dog (1 of 426) and cat (1 of 215) [35], and SFTSV infection was recently reported in a 50-year-old woman from Japan who died after being bitten by a stray cat [36].
In addition, wild animals may also be SFTSV reservoirs. For example, in Korea the Korean wild deer (KWD, n=21), Siberian roe deer (n=3), goral (n=5), raccoon dog (n=7), wild boar (WB, n=54), and carrion cow (n=1) were surveyed for SFTSV and one KWD (4.76%) and two WB (3.70%) were found to be positive by reverse transcription polymerase chain reaction (RT-PCR) and nested nested polymerase chain reaction (Nested PCR) [37]. In China, SFTSV was also detected in small wild animals during the tick-feeding season with SFTSV-RNA being detected in hedgehogs (5/93, 5.4%), common shrews (2/102, 2.0%), fulvous harvest mice (1/46, 2.2%), and house mice (1/168, 0.6%). Moreover, the presence of SFTS viral antibodies was highest in hedgehogs (13/93, 14.0%) [37]. Furthermore, in six areas of Japan wild boars were assessed for SFTSV antibodies with 51% of samples from Nagasaki (27/53), 25% from Omura (1/4), and 19% from Matsuura (7/37) being positive [38].
While rodents are known to be reservoir hosts of Bunyaviruses, previous surveillance studies of SFTSV prevalence have yielded contradictory results. For example, one study reported that the SFTSV antigen rate was only 4.31% (76/1,762) in rats (detected in heart, liver, spleen, lung, kidney, and brain) from 18 provinces in China while it was 11.4% (8/70) in Zhejang Province [3940]. In Jiangsu China, only 4.4% seroprevalence for SFTSV was detected in rodents [36]. Further, in Zheijang Province, China, two of 52 rodents captured (striped field mice) tested positive for SFTSV RNA in lung tissue and this virus exhibited a high level of sequence homology with virus isolated from human cases in the nearby village [41]. However, in a study done in Beijing, China, no SFTSV RNA was detected in sera from 81 rodents [42]. Thus, although seropositivity has been reported in rodents, more study is required as well as further examination of the role of rodents in the transmission of SFTSV to other animals and humans [30].
Migratory wild birds are also known to be transmitters of infectious diseases including influenza, Lyme disease, and West Nile virus [43]. Because wild birds often travel long distances to seek food and for breeding purposes and because they carry different parasites, including ticks, they can disseminate diseases including SFTS [4344]. Based on several studies, SFTS are believed to be transferred from one place to other by migratory birds since H. longicornis which serves as transmitters of SFTSV are regular parasites of migratory birds that are distributed in Asia-pacific region. Furthermore, the geographic distribution of these ticks overlaps with the migratory bird flyways in China, South Korea, and Japan [11428]. Additionally, phylogenetic analysis of SFTSVs isolated in South Korea revealed it to be similar to samples from Japan and Korea [45]. One tick surveillance study conducted on birds from Jeju Island in South Korea, found H. longicornis on several species of migratory birds including Zoothera aurea, Turdus hortulorum, Halcyon coromanda, and Pitta nympha [46]. Furthermore, SFTSV infection was reported to result in fatalities in 10 out of 36 human patients on Jeju Island suggesting this is a high-prevalence region [47] and thereby suggesting migratory birds may be involved in the transmission of SFTS on the island. Therefore, more detailed studies regarding the role of migratory birds in SFTS transmission are required.
Compared to the number of human case studies and surveillance conducted in China, South Korea, and Japan, a limited number of animal studies have been conducted largely due to the rarity of SFTS clinical symptoms in animals [32]. However, the seropositive status of many different animals may suggest that these domesticated and wild animals act as amplifying hosts of SFTSV and play a major role in feeding the infected ticks which can later spread the SFTS infection. Therefore, further studies are needed to fully understand the capacity of SFTSV to infect animals.
Go to :
Control Options for SFTSV Infections
Because SFTSV is a relatively novel disease, therapeutic options are still limited; administration of ribavirin, steroids, and/or plasma exchange in human patients [48]. Since the discovery of SFTS in 2009 in China [1], several companies and laboratories have been working to develop vaccines for SFTSV [49]. However, there are many hurdles which must be overcome to successfully produce an effective vaccine including the establishment of reliable vaccine efficacy testing methods. Recently, serum neutralizing antibody-based testing methods were demonstrated; however, the cross-reactivity of these antibodies with different SFTSV genotypes still needs to be evaluated to define a standard strain for vaccine production [47]. Further, the establishment of proper animal infection models will be necessary to evaluate vaccine efficacy in vivo. To date, only a few animal models such as the type I interferon (IFN)-deficient mouse model [5051], newborn mice [52], and mitomycin-treated mice [53] (Table 2) have been used to recapitulate the fatal illness following SFTSV infection. It should be noted that the type I IFN-deficient and mitomycin-treated mouse models are both immunosuppressed models. This means they cannot deliver the normal immune response against virus infection, and hence, a model of SFTSV infection with normal immune status will be necessary in order to test vaccine efficacy.
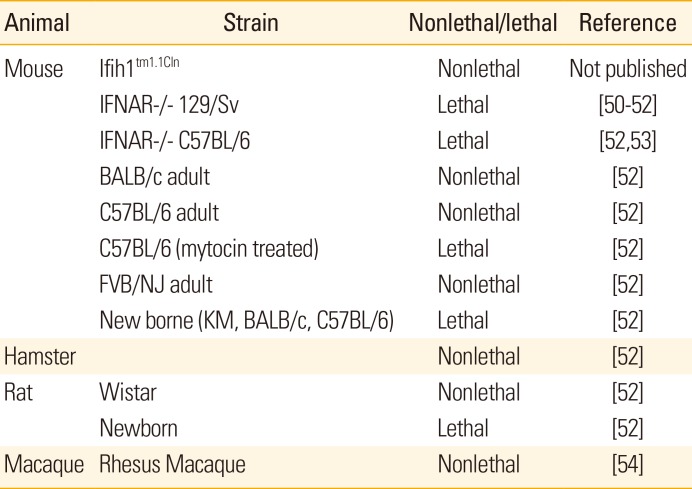
Table 2
Animal models for SFTSV infection
| Animal | Strain | Nonlethal/lethal | Reference |
|---|---|---|---|
| Mouse | Ifih1tm1.1Cln | Nonlethal | Not published |
| IFNAR-/- 129/Sv | Lethal | [50,51,52] | |
| IFNAR-/- C57BL/6 | Lethal | [52,53] | |
| BALB/c adult | Nonlethal | [52] | |
| C57BL/6 adult | Nonlethal | [52] | |
| C57BL/6 (mytocin treated) | Lethal | [52] | |
| FVB/NJ adult | Nonlethal | [52] | |
| New borne (KM, BALB/c, C57BL/6) | Lethal | [52] | |
| Hamster | Nonlethal | [52] | |
| Rat | Wistar | Nonlethal | [52] |
| Newborn | Lethal | [52] | |
| Macaque | Rhesus Macaque | Nonlethal | [54] |
![]()
Studies of the T-705 (Favipiravir) therapeutic in Japan suggest that this drug may be a good candidate for the treatment of SFTSV infection. Intraperitoneal or oral administration of this drug resulted in 100% survivability of IFNAR−/− mice infected with a lethal dose of SFTSV without weight loss and with a reduced viral load in the serum [51]. Although Favipiravir was found to have promising results in this mouse model, detailed studies in other animal models will be needed to prove its efficacy and safety.
Go to :
Conclusion
SFTSV poses an imminent threat to public health due to its wide range of animal hosts and the ability to cause severe infections in humans, especially in the elderly. Continuous monitoring for and molecular detection of this virus are important for correct diagnosis of SFTSV infection. Because animals do not show overt clinical symptoms upon infection and clinical symptoms of SFTSV infection resemble other diseases in humans, it is possible this virus is already present in other countries aside from China, Korea, and Japan. Although there have been several studies focused on the pathogenesis and transmission of this virus, there is currently no vaccine available to prevent the spread of SFTSV. Initial studies indicate a higher mortality rate in patients 50 years and above suggesting susceptibility to SFTS infection may vary depending on several risk factors including patient age and immune status. Nonetheless, with initial indications in China, Korea, and Japan of an overall mortality rate in the range of 30%–40%, and a wide range of host species that display limited clinical illnesses and have direct human contact, vaccine development will be critical for the prevention and eradication of SFTSV.
Go to :
 XML Download
XML Download