

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Toxoplasma gondii belongs to the Apicomplexa phylum that caused a widespread zoonotic infection in a wide range of intermediate hosts, including men, livestock, birds, marine mammals, and snakes [1234567]. More than one century ago, this protist was discovered from the spleen and liver smears of a north African rodent namely Ctenodactylus gondii [8]. Recent review papers have documented over one-third of peolple throughout the globe are latently infected with T. gondii and carry it [7910].
The biologic stages and the main transmission routes of T. gondii are depicted in Fig. 1. Briefly, Toxoplasma has three infectious stages: sporozoites (in oocysts), tachyzoites (rapidly multiplying form), and bradyzoites (tissue cyst form). Tachyzoites as rapidly multiplying forms are the main reason of the acute phase of toxoplasmosis and responsible for a wide spectrum of clinical signs. Once a host becomes infected, the parasite can survive with complex mechanisms for the whole lifespan within tissue cysts located usually in the skeletal muscles, brain, eyes, and myocardium. In some circumstances, particularly upon the suppression of immune system, latent encysted parasites can reactivate and the symptoms of infection became evident [1112].
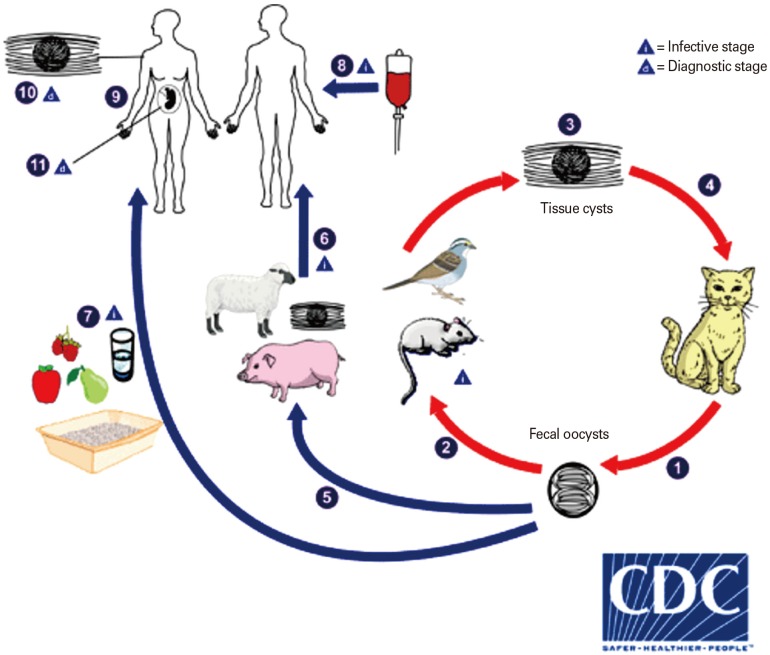 | Fig. 1The biologic stages and the main transmission routes of Toxoplasma gondii. The only known definitive hosts for T. gondii are members of family Felidae (domestic cats and their relatives). Unsporulated oocysts are shed in the cat's feces (1). Although oocysts are usually only shed for 1–2 weeks, large numbers may be shed. Oocysts take 1–5 days to sporulate in the environment and become infective. Intermediate hosts in nature (including birds and rodents) become infected after ingesting soil, water, or plant material contaminated with oocysts (2). Oocysts transform into tachyzoites shortly after ingestion. These tachyzoites localize in neural and muscle tissue and develop into tissue cyst bradyzoites (3). Cats become infected after consuming intermediate hosts harboring tissue cysts (4). Cats may also become infected directly by ingestion of sporulated oocysts. Animals bred for human consumption and wild game may also become infected with tissue cysts after ingestion of sporulated oocysts in the environment (5). Humans can become infected by any of several routes: eating undercooked meat of animals harboring tissue cysts (6); consuming food or water contaminated with cat feces or by contaminated environmental samples (such as fecal-contaminated soil or changing the litter box of a pet cat) (7); blood transfusion or organ transplantation (8); transplacentally from mother to fetus (9). Diagnosis is usually achieved by serology, although tissue cysts may be observed in stained biopsy specimens (10). Diagnosis of congenital infections can be achieved by detecting T. gondii DNA in amniotic fluid using molecular methods such as polymerase chain reaction (11) Adapted from Centers for Disease Control and Prevention [20].
|
Toxoplasmosis predominantly is asymptomatic in immunocompetent persons, while may cause severe complications with life-threatening outcomes in immunosuppressed persons such as human immunodeficiency virus/acquired immune deficiency syndrome subjects, patients with malignancy and transplant recipients [310131415]. Upon maternal infection, fetus is probably to be exposed with vertical transmission, which may cause abortion in those pregnant women that acquired toxoplasmosis during her pregnancy. Notably, the complications will be differ according to the gestational age ranged from retardation, microcephaly, hydrocephalus, brain focal lesions, deafness, so on [16171819]. In the other hand, this unicellular protozoan parasite may result in fetal death, neonatal loss, and abortion in domestic animals, especially in goats and sheep which imposes heavy economic losses in the industry of veterinary medicine and animal husbandry. Furthermore, they serve as a source of transmission to humans [321]. Interestingly, it has been shown chronic toxoplasmosis considerably associated with some autoimmune diseases, mental and neurodegenerative disorders. In addition, the scientists frequently have confirmed that T. gondii infection can manipulate and alter the behavior, not only in animal models, but in men as well [2223242526].
Go to : 
Vaccine Candidate: Current State and Future
The present common primary control measures for men and animals T. gondii infection depends on chemotherapy. The drugs are not sufficiently satisfactory due to their side effects. For example, many concerns remain regarding the use of these medications in pregnant women because of the probable teratogenic effect on the fetus [27]. Besides, the routinely prescribed therapeutic medicines for the treatment of toxoplasmosis, can limit the proliferation of tachyzoites in the onset of infection, while they are unable to eliminate them and have no effect on the encysted parasites within infected hosts [27]. Hence, discover and development of an effective vaccine has high priority and urgently needed to prevent and control toxoplasmosis, because of the heavy burden of chronic infection throughout the world in different host species, particularly in men and domestic animals. For this reasons, several vaccine types with different approaches have been experimentally evaluated worldwide [282930313233343536].
During three recent decades, continous efforts of researchers have made precious successes in the development of T. gondii candidate vaccines mainly on dense granule antigens, microneme antigens, rhoptry antigens (ROP), and surface antigens. These vaccines predominantly induced partial protection along with reduction of brain cyst load post challenge with virulent and/or avirulent T. gondii strains in mouse models [3031323337]. Currently, there is a commercially licensed vaccine known as Toxovax (the live-attenuated tachyzoites of T. gondii S48 strain) which used in the veterinary industry in some countries [38]. Toxovax reduces the incidence of congenital toxopllasmosis in sheep; however, it is not useful for human vaccination, particularly in immunosuppressed subjects, because of its reversion to the wild-type virulence as previously the accidental infections have been reported in farmers [283133].
In regard to the major transmission routes of parasite and the toxoplasmosis manifestations in high risk groups, the main targets for vaccination strategy would cover [2939]:
-
‒ Vaccination to prevent acute parasitemia and protect against congenital toxoplasmosis
‒ Prevent or reduce tissue cysts in food animals to interrupt the transmission route to humans‒ Prevention or reduction of oocyst shedding in cats to confine environmental contamination as well as minimize the risk of toxoplasmosis for all intermediate hosts
Go to :
Calcium-Dependent Protein Kinase
The increasing number of experiments over the past decade has focused on the evaluation of the immunogenicity of main functional proteins including kinases and enzymes. These functional proteins, mainly involve in the duplication, transcription, metabolism, signaling pathways and other biological procedures of the parasite life cycle [4041424344454647]. Based on genomic analysils, T. gondii encoded 108 protein kinases (presumably with a catalytic activity) and 51 pseudokinases genes (due to the absence of a catalytic domain) [48]. As signaling mediators of calcium-related signaling cascades, the calcium-dependent protein kinases (CDPKs) are a distinct protein kinase family, which belongs to the superfamily of kinases [495051]. CDPKs are restricted to the apicomplexans, ciliates, and plants. In apicomplexan, particularly T. gondii, several CDPKs (as the most crucial household proteins) have been characterized involving in a different array of functions in the life cycle of parasite, including gliding motility, cell invasion and egress as well as some other pivotal developmental processes [495152]. CDPKs not expressed by mammals and fungi which indicates validated targets that can be exploited for a promising target vaccine candidate against toxoplasmosis and anti-T. gondii drugs [5253545556]. Out of 14 CDPKs present in T. gondii, CDPK1, CDPK3, CDPK4, CDPK5, CDPK6, and CDPK7 are the only CDPKs which expressed and conserved in almost all apicomplexan parasites [495758]. The specific features and main functions of some CDPKs have been embedded in Table 1.
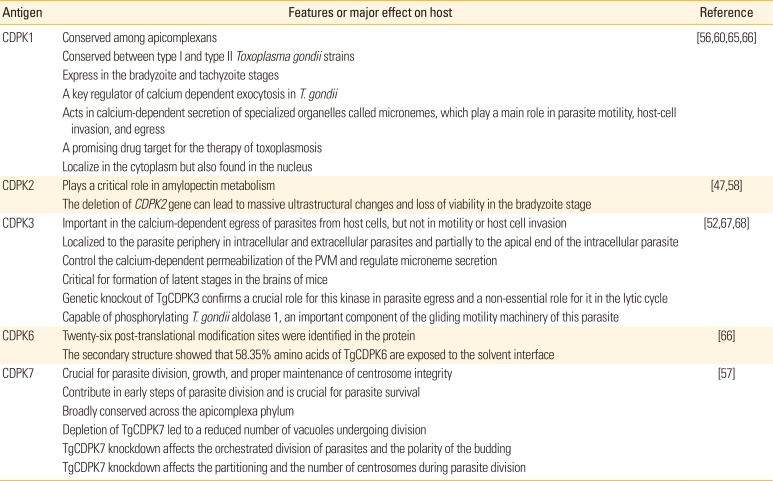
Table 1
The main features and functions of some CDPKs
| Antigen | Features or major effect on host | Reference |
|---|---|---|
| CDPK1 | Conserved among apicomplexans | [56,60,65,66] |
| Conserved between type I and type II Toxoplasma gondii strains | ||
| Express in the bradyzoite and tachyzoite stages | ||
| A key regulator of calcium dependent exocytosis in T. gondii | ||
| Acts in calcium-dependent secretion of specialized organelles called micronemes, which play a main role in parasite motility, host-cell invasion, and egress | ||
| A promising drug target for the therapy of toxoplasmosis | ||
| Localize in the cytoplasm but also found in the nucleus | ||
| CDPK2 | Plays a critical role in amylopectin metabolism | [47,58] |
| The deletion of CDPK2 gene can lead to massive ultrastructural changes and loss of viability in the bradyzoite stage | ||
| CDPK3 | Important in the calcium-dependent egress of parasites from host cells, but not in motility or host cell invasion | [52,67,68] |
| Localized to the parasite periphery in intracellular and extracellular parasites and partially to the apical end of the intracellular parasite | ||
| Control the calcium-dependent permeabilization of the PVM and regulate microneme secretion | ||
| Critical for formation of latent stages in the brains of mice | ||
| Genetic knockout of TgCDPK3 confirms a crucial role for this kinase in parasite egress and a non-essential role for it in the lytic cycle | ||
| Capable of phosphorylating T. gondii aldolase 1, an important component of the gliding motility machinery of this parasite | ||
| CDPK6 | Twenty-six post-translational modification sites were identified in the protein | [66] |
| The secondary structure showed that 58.35% amino acids of TgCDPK6 are exposed to the solvent interface | ||
| CDPK7 | Crucial for parasite division, growth, and proper maintenance of centrosome integrity | [57] |
| Contribute in early steps of parasite division and is crucial for parasite survival | ||
| Broadly conserved across the apicomplexa phylum | ||
| Depletion of TgCDPK7 led to a reduced number of vacuoles undergoing division | ||
| TgCDPK7 knockdown affects the orchestrated division of parasites and the polarity of the budding | ||
| TgCDPK7 knockdown affects the partitioning and the number of centrosomes during parasite division |
![]()
It is well known that gene knockouts of CDPKs may affect on specific developmental stages [49]. For instance, Billker et al. [59] showed the disruption of CDPK4 in asexual stages of Plasmodium berghei leads to differentiation defects in male gametocytes. They concluded “CDPK4 is essential for the sexual reproduction and mosquito transmission of P. berghei.” Furthermore, it has been shown conditional suppression of CDPK1 protein lead to loss of some capabilities, including parasite motility, host cell invasion, and egress [60]. The T. gondii kinase CDPK2 has functional Ca2+ and carbohydrate-binding domains, which its deficiency causes hyperaccumulation of starch in Toxoplasma parasites results death of chronic-stage parasites [47]. In another study, Morlon-Guyot et al. [57] reported that the downregulation of CDPK7 protein in T. gondii results in pronounced defects in parasite division and a major growth deficiency. The polarity of daughter cells budding and the fate of several subcellular structures or proteins involved in cell division were affected, as well. The authors have remarked “CDPK7 is crucial for proper maintenance of centrosome integrity required for the initiation of endodyogeny” [57]. More recently, Wang and colleagues assessed the functions of six different CDPKs including CDPK4, CDPK4A, CDPK5, CDPK6, CDPK8, and CDPK9 in T. gondii to determine are they suitable for designing as drug targets or not? For this purpose, clustered regularly interspaced short palindromic repeats/Cas9 system was employed to disrupt the CDPKs genes by insertion of dihydrofolate reductase and ultimately the six knockout (KO)-CDPK strains were successfully identified by indirect immunofluorescence. The findings revealed a lack of considerable difference between the six KO-CDPK strains and wild-type strain in virulence and the lytic cycle in terms of invasion, egress, and replication. The authors have highlighted “these CDPKs seems not pivotal for parasite lytic cycle and also not virulence factors for mice, probably they contribute in other functions of parasite” [61].
Go to :
CDPK DNA Vaccine
DNA vaccination as a robust method have become a major focus with many advantages compared to traditional vaccines in several parameters as follows [626364]:
-
- Design (more rapid design as well as can be rapidly isolated and cloned)
- Versatility (ease in adapting or improving plasmid sequence, capability to deliver multi-antigen vaccines into a host only with a single dose, ease in formulation with different adjuvants)- Production (cost effective, ease of production, capable of large-scale production, appropriate protein folding for correct epitope expression)- Transport (stability at room temperature and no need to cold chain)- Safety (cannot revert to the pathogenic form, safer than live or attenuated vaccines)- Immune responses (boost the expression of an encoded vaccine antigen lwithin host cells, able to induce a longlasting immunity, elicits efficient and specific humoral and cellular immune responses, provide immune priming but poor immune boosting)
DNA vaccination recruits the plasmid vector in order to transfer and expression of the target gene and strongly elicit specific humoral and cell-mediated immune responses [3462]. After the inoculation, the DNA enter to the cell cytoplasm then express the encoded protein target inside the host cell [62]. In general, during the course of toxplasmosis both cell-mediated and humoral immune responses are elicited, so that the secretion of anti‒T. gondii specific antibodies hamper the attachment of parasites to related host cell receptors. Furthermore, the specific-IgG antibodies have a critical role in restriction of toxoplasmosis through activation of the classical complement cascades, enhancement of killing activity of macrophages to eliminate the intracellular parasites and other roles against this opportunistic agent [69]. Besides, the production of interferon-γ (IFN-γ) as the adaptive cellular immunity, limit the growth of T. gondii in different infection stages [7071]. Notably, protection against T. gondii infection predominantly is developed through both types of CD8+ and CD4+ T cells as cell-mediated immunity arms. Although, as it is well established CD8+ T cell and IFN-γ are obvious to be more fundamental to restrict toxoplasmosis [33]. Recently, was shown that DNA vaccination with CDPK1, CDPK2, CDPK3, and CDPK5 dramatically increased the number of CD4+ and/or CD8+ T cells in immunized mice [5254555658].
There are several limitations affecting the immune efficacy of DNA vaccines, which occasionally confined the immunogenicity of them such as dosages of inoculum and the delivery route [62]; however, 100 µg of the plasmid are routinely injected intramuscularly in mice [5254555658]. It should be noted, following the inoculation, the naked DNA poorly distributed and rapidly degraded by lysosomes and DNAses, thereby, reduce the expression of plasmid DNA [727374]. Thus, the use of adjuvants as a robust strategy has become popular because of the following reasons [62637475]:
-
- Augment the immunogenicity of DNA vaccines
- Boost DNA delivery- Increased the magnitude/duration of plasmid DNA expression- Recruit the immune cells to the site of inoculation, thereby increases the immunostimulatory features of plasmid- Help the uptake of DNA into host cells or by professional antigen-presenting cells (APCs)- Protect plasmid against degradation by DNAses
An increasing number of papers have demonstrated that the use of genetic adjuvants such as cytokines (IFN-γ, interleukin [IL]-7, IL-12, IL-15, IL18, IL-21, etc.), chemokines, costimulatory molecules (B7-1, B7-2, etc.), etc. strongly augment the protective efficacy of DNA vaccine [3334545676777879]. Genetic adjuvants can be either encoded on a separate vector, or expressed on the same vector as the antigen and thereafter co-administrated with the vaccine [34626375].
CDPK1 as an essential regulator of calcium-dependent exocytosis in T. gondii is conserved between both type I and type II T. gondii strains which expressed in the bradyzoite and tachyzoite stages [5660]. TgCDPK1 was localized in the cytoplasm, but also found in the nucleus [65]. It has been shown that Kunming mice immunized with pVAX-CDPK1 survived for a significant longer time post intraperitoneally challenge with 1×103 tachyzoites of RH strain. Also, the immunized mice showed a mixed Th1/Th2 response with the predominance of IgG2a levels over IgG1, increased secretion of IFN-γ, IL-2, IL-4, and IL-10 cytokines, enhanced splenocyte proliferation and percentages of CD4+ and CD8+ T cells as well as reduction of brain cyst load, compared with control groups (p<0.05) [5456]. Notably, co-delivery of pVAX/IL-21/IL-15 or pVAX/IL-7/IL-15 plus pVAX-CDPK1 considerably boosted the above-mentioned outcomes in mice (p<0.05) [5456]. The co-injection of plasmids encoding cytokine is one the best approaches to boost the efficiency of DNA vaccines that has several advantages, including ease of cloning process, simplicity, and cost benefits [63]. For instance, IL-7 and IL-15 are dominantly involved in the generation and homeostasis of memory T cells [80]. Besides, it is well established that signaling by both IL-7 and IL-15 cytokines could synergize to boost the generation of memory T cells [81]. As common g chain cytokines, including IL-2, IL-4, IL-7, IL-9, IL-15, and IL-21, recent papers have reported co-administration of pVAX/IL-21/IL-15 plus pVAX-MIC8 or pVAX-CDPK1 promoted protective immunity of DNA vaccines expressing MIC8 and CDPK1 [5682]. More examples of immunization experiments with DNA vaccines against T. gondii have been listed in Table 2.
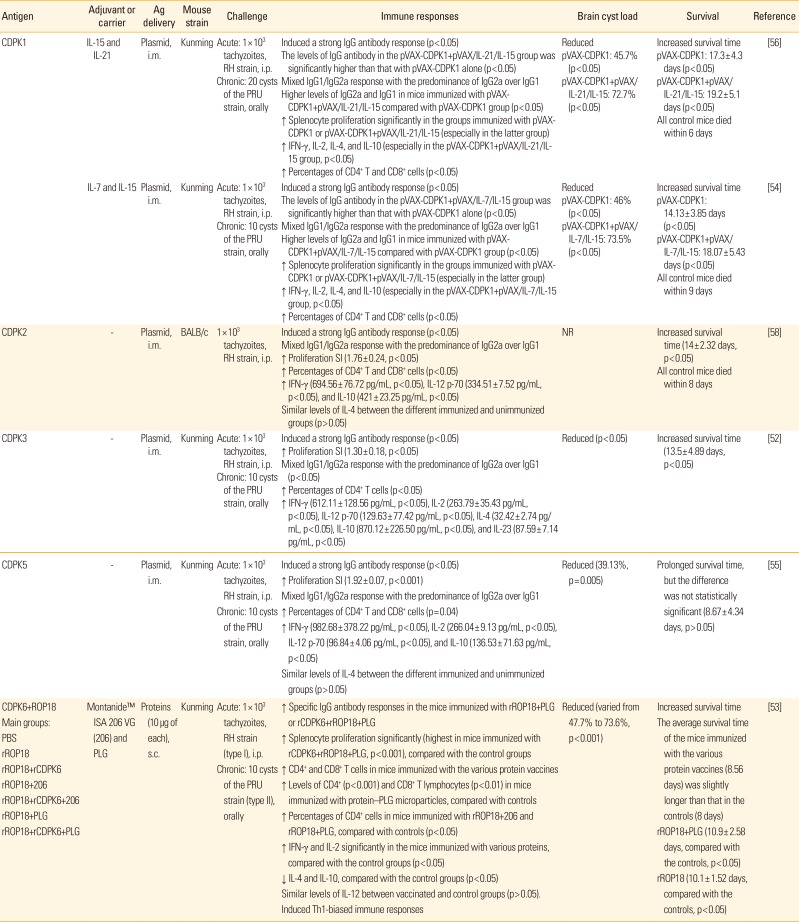
Table 2
Baseline characteristics of included studies based on immunization experiments with DNA or protein vaccines against Toxoplasma gondii in mouse models
| Antigen | Adjuvant or carrier | Ag delivery | Mouse strain | Challenge | Immune responses | Brain cyst load | Survival | Reference |
|---|---|---|---|---|---|---|---|---|
| CDPK1 | IL-15 and IL-21 | Plasmid, i.m. | Kunming | Acute: 1×103 tachyzoites, RH strain, i.p. | Induced a strong IgG antibody response (p<0.05) | Reduced | Increased survival time | [56] |
| Chronic: 20 cysts of the PRU strain, orally | The levels of IgG antibody in the pVAX-CDPK1+pVAX/IL-21/IL-15 group was significantly higher than that with pVAX-CDPK1 alone (p<0.05) | pVAX-CDPK1: 45.7% (p<0.05) | pVAX-CDPK1: 17.3± 4.3 days (p<0.05) | |||||
| Mixed IgG1/IgG2a response with the predominance of IgG2a over IgG1 | pVAX-CDPK1+pVAX/IL-21/IL-15: 72.7% (p<0.05) | pVAX-CDPK1+pVAX/IL-21/IL-15: 19.2± 5.1 days (p<0.05) | ||||||
| Higher levels of IgG2a and IgG1 in mice immunized with pVAX-CDPK1+pVAX/IL-21/IL-15 compared with pVAX-CDPK1 group (p<0.05) | All control mice died within 6 days | |||||||
| ↑ Splenocyte proliferation significantly in the groups immunized with pVAX-CDPK1 or pVAX-CDPK1+pVAX/IL-21/IL-15 (especially in the latter group) | ||||||||
| ↑ IFN-γ, IL-2, IL-4, and IL-10 (especially in the pVAX-CDPK1+pVAX/IL-21/IL-15 group, p<0.05) | ||||||||
| ↑ Percentages of CD4+ T and CD8+ cells (p<0.05) | ||||||||
| IL-7 and IL-15 | Plasmid, i.m. | Kunming | Acute: 1×103 tachyzoites, RH strain, i.p. | Induced a strong IgG antibody response (p<0.05) | Reduced | Increased survival time | [54] | |
| Chronic: 10 cysts of the PRU strain, orally | The levels of IgG antibody in the pVAX-CDPK1+pVAX/IL-7/IL-15 group was significantly higher than that with pVAX-CDPK1 alone (p<0.05) | pVAX-CDPK1: 46% (p<0.05) | pVAX-CDPK1: 14.13±3.85 days (p<0.05) | |||||
| Mixed IgG1/IgG2a response with the predominance of IgG2a over IgG1 | pVAX-CDPK1+pVAX/IL-7/IL-15: 73.5% (p<0.05) | pVAX-CDPK1+pVAX/IL-7/IL-15: 18.07± 5.43 days (p<0.05) | ||||||
| Higher levels of IgG2a and IgG1 in mice immunized with pVAX-CDPK1+pVAX/IL-7/IL-15 compared with pVAX-CDPK1 group (p<0.05) | All control mice died within 9 days | |||||||
| ↑ Splenocyte proliferation significantly in the groups immunized with pVAX-CDPK1 or pVAX-CDPK1+pVAX/IL-7/IL-15 (especially in the latter group) | ||||||||
| ↑ IFN-γ, IL-2, IL-4, and IL-10 (especially in the pVAX-CDPK1+pVAX/IL-7/IL-15 group, p<0.05) | ||||||||
| ↑ Percentages of CD4+ T and CD8+ cells (p<0.05) | ||||||||
| CDPK2 | - | Plasmid, i.m. | BALB/c | 1×103 tachyzoites, RH strain, i.p. | Induced a strong IgG antibody response (p<0.05) | NR | Increased survival time (14±2.32 days, p<0.05) | [58] |
| Mixed IgG1/IgG2a response with the predominance of IgG2a over IgG1 | All control mice died within 8 days | |||||||
| ↑ Proliferation SI (1.76±0.24, p<0.05) | ||||||||
| ↑ Percentages of CD4+ T and CD8+ cells (p<0.05) | ||||||||
| ↑ IFN-γ (694.56±76.72 pg/mL, p<0.05), IL-12 p-70 (334.51±7.52 pg/mL, p<0.05), and IL-10 (421±23.25 pg/mL, p<0.05) | ||||||||
| Similar levels of IL-4 between the different immunized and unimmunized groups (p>0.05) | ||||||||
| CDPK3 | - | Plasmid, i.m. | Kunming | Acute: 1×103 tachyzoites, RH strain, i.p. | Induced a strong IgG antibody response (p<0.05) | Reduced (p<0.05) | Increased survival time (13.5±4.89 days, p<0.05) | [52] |
| Chronic: 10 cysts of the PRU strain, orally | ↑ Proliferation SI (1.30±0.18, p<0.05) | |||||||
| Mixed IgG1/IgG2a response with the predominance of IgG2a over IgG1 (p<0.05) | ||||||||
| ↑ Percentages of CD4+ T cells (p<0.05) | ||||||||
| ↑ IFN-γ (612.11±128.56 pg/mL, p<0.05), IL-2 (263.79±35.43 pg/mL, p<0.05), IL-12 p-70 (129.63±77.42 pg/mL, p<0.05), IL-4 (32.42±2.74 pg/mL, p<0.05), IL-10 (870.12± 226.50 pg/mL, p<0.05), and IL-23 (87.59±7.14 pg/mL, p<0.05) | ||||||||
| CDPK5 | - | Plasmid, i.m. | Kunming | Acute: 1×103 tachyzoites, RH strain, i.p. | Induced a strong IgG antibody response (p<0.05) | Reduced (39.13%, p=0.005) | Prolonged survival time, but the difference was not statistically significant (8.67±4.34 days, p>0.05) | [55] |
| Chronic: 10 cysts of the PRU strain, orally | ↑ Proliferation SI (1.92±0.07, p<0.001) | |||||||
| Mixed IgG1/IgG2a response with the predominance of IgG2a over IgG1 | ||||||||
| ↑ Percentages of CD4+ T and CD8+ cells (p=0.04) | ||||||||
| ↑ IFN-γ (982.68±378.22 pg/mL, p<0.05), IL-2 (266.04±9.13 pg/mL, p<0.05), IL-12 p-70 (96.84±4.06 pg/mL, p<0.05), and IL-10 (136.53±71.63 pg/mL, p<0.05) | ||||||||
| Similar levels of IL-4 between the different immunized and unimmunized groups (p>0.05) | ||||||||
| CDPK6+ROP18 | Montanide™ ISA 206 VG (206) and PLG | Proteins (10 µg of each), s.c. | Kunming | Acute: 1×103 tachyzoites, RH strain (type I), i.p. | ↑ Specific IgG antibody responses in the mice immunized with rROP18+PLG or rCDPK6+rROP18+PLG | Reduced (varied from 47.7% to 73.6%, p<0.001) | Increased survival time | [53] |
|
Main groups: PBS rROP18 rROP18+rCDPK6 rROP18+206 rROP18+rCDPK6+206 rROP18+PLG rROP18+rCDPK6+PLG |
Chronic: 10 cysts of the PRU strain (type II), orally | ↑ Splenocyte proliferation significantly (highest in mice immunized with rCDPK6+rROP18+PLG, p<0.001), compared with the control groups | The average survival time of the mice immunized with the various protein vaccines (8.56 days) was slightly longer than that in the controls (8 days) | |||||
| ↑ CD4+ and CD8+ T cells in mice immunized with the various protein vaccines | rROP18+PLG (10.9±2.58 days, compared with the controls, p<0.05) | |||||||
| ↑ Levels of CD4+ (p<0.001) and CD8+ T lymphocytes (p<0.01) in mice immunized with protein–PLG microparticles, compared with controls | rROP18 (10.1±1.52 days, compared with the controls, p<0.05) | |||||||
| ↑ Percentages of CD4+ cells in mice immunized with rROP18+206 and rROP18+PLG, compared with controls (p<0.05) | ||||||||
| ↑ IFN-γ and IL-2 significantly in the mice immunized with various proteins, compared with the control groups (p<0.05) | ||||||||
| ↓ IL-4 and IL-10, compared with the control groups (p<0.05) | ||||||||
| Similar levels of IL-12 between vaccinated and control groups (p>0.05). | ||||||||
| Induced Th1-biased immune responses |
![]()
Go to :
Recombinant CDPK Protein Vaccine
During recent decades, continuous attempts of researchers have revealed great achievements to clarify the different perspectives of the cell and molecular biology of T. gondii [495760676883848586]. These efforts resulted great motivation in the current focus on vaccine development against chronic and acute toxoplasmosis, based on the distinctive subcellular components of the parasite [282930313234]. Recombinant subunit vaccines provide an alternative way for the development of vaccine candidates in both animals and men. These vaccine types can induce humoral and cellular immune responses as well as they are very suitable for large-scale production [3031]. Application of traditional and genetic adjuvants has become popular, due to their potential capability in augmenting specific and long-lasting protective immunity [7587].
Owing to proteolytically degradation of recombinant proteins, presence of a proper delivery system is necessary to protect them from degradation [88]. Polylactide-co-glycolide (PLG) is a biodegradable and biocompatible polymer which extend the protein releasing period to induce a long-lasting immune response as well as reduce the protein degradation. Besides, PLG by increasing the uptake of antigen and its presentation by APCs would augment the protective immunity. This biodegradable polymer encapsulates the recombinant subunit vaccines, which maintain their antigenicity to boost the efficacy [5389]. In this context, Zhang et al. (2016) [53] designed a comprehensive study on two recombinant proteins rROP18 and rCDPK6 adjuvanted with PLG and Montanide ISA 206 VG (206). In brief, the mice were vaccinated subcutaneously with different proteins (with or without adjuvant) and then challenged intraperitoneally with 1×103 tachyzoites of RH strain (type I) and orally with 10 cysts of the PRU strain (type II) for acute and chronic infection, respectively. The findings revealed both recombinant protein vaccines promoted specific humoral and cellular Th1-biased immune responses, high production of IFN-γ and IL-2 cytokines with strong lymphocyte proliferative responses. The titers of IgG values in Kunming mice vaccinated with rROP18+PLG or rCDPK6+rROP18+PLG were dramatically higher than rROP18 or rCDPK6+rROP18 (p<0.01) groups, however, were not dramatically different from those mice injected with various proteins-montanide 206 adjuvant (p>0.05). Also, the highest splenocyte proliferative responses were observed in the mice vaccinated with rROP18+rCDPK6+PLG (p<0.001). Moreover, increased percentage of CD4+ and CD8+ T cells were recorded in mice immunized with the various protein vaccines (especially in protein–PLG groups). The brain tissue cyst burden in mice immunized with the various proteins was significantly reduced than those in control groups (p<0.001), ranging from 47.7% to 73.6%. The authors concluded that “the use of PLG is superior than Montanide 206, because of encapsulated proteins into PLG conferred immunity to T. gondii for an extended period. However, evaluation of the potentcy of PLG in the development of a vaccine against toxoplasmosis should be further studied in future” [53]. More details can be found in Table 2. Application of PLG on other antigens frequently have been tested and showed acceptable outcomes [909192]. For instance, rROP18 encapsulated in nanoparticles with polylactide-co-glycolide acid (rROP18+PLGA), significantly increased the values of IgG antibody, elicited Th1-type responses with the predominance of IgG2a over IgG1, and enhanced intestinal IgA values [91]. In another investigation, rROP18 and rROP38 encapsulated into PLG elicited both humoral and cell-mediated immune responses in Kunming mice in terms of total IgG titers (p<0.01), IgG2a subclass (p<0.01), IFN-γ cytokine (p<0.01) and mixed Th1/Th2 immunity responses (but bias to Th1). Also, the formulation of mixed antigens (rROP38+rROP18+PLG) boosted the efficacy of vaccine [90]. The above findings suggest that PLG could be a promising novel adjuvant with preserving the protein immunogenicity for longer duration.
Go to :
CDPK Peptide Vaccine
More recently, bioinformatics as a novel interdisciplinary science has become favorite which analyze the biological data using defined technologies and algorithms from computer sciences, mathematics, statistics, physics, biology, and medicine [93]. This new science has several advantages in comparison to traditional methods such as satisfactory precision and accuracy, relatively inexpensive and affordable, etc. Nowadays, Bioinformatics is extensively used to predict protein structures, biological characteristics, functions, epitopes, and design of new vaccines [939495]. The prediction of epitopes is highly fundamental to assess the immunogenicity of an antigen in design of reverse vaccines. Accordingly, bioinformatics softwares and online databases could help scientists to predict and recognize the potential B and T cell epitopes [939596]. In this regards, Zhang et al. (2014) [66] designed an investigation using multiple Bioinformatics approaches on TgCDPK6 protein to identify potential antigenicity, signal peptides, topological structures, epitopes, and other chemical and physical characteristics. They showed 26 post-translational modification sites in the protein. In addition, the secondary structure of CDPK6 showed that nearly 58% of its amino acids are exposed to the solvent interface. The prediction of functional domains revealed TgCDPK6 possess seven protein kinase C phosphorylation sites and one protein kinase adenosine triphosphate (ATP)-binding region. In the other hand, the high hydrophilic domains were distributed in amino acid positions 21–59, 68–81, 156–205, 245–271, 280–294, 297–324, 334–356, 367–393, 474–498, and 543–553. The advanced structure of TgCDPK6 was developed by a homology modeling method and then validated by PROCHECK, represented that most amino acid residues were in the most favored regions. In the study, ten potential epitopes were recognized. The authors have concluded that “TgCDPK6 could be a highly potent DNA vaccine candidate against T. gondii; however, more experimentally studies are needed in future” [66]. It should be noted, a recent study by the same authors from the same lab demonstrated satisfactory outcomes following vaccination of Kunming mice with the formulation of rCDPK6+rROP18+PLG [53].
There is a lack of study regarding epitope-based vaccines on CDPKs. Previously was shown that multi-epitope-based vaccines, strongly elicited both humoral and cellular immune responses, thereby significantly prolonged survival time along with reduction of brain cyst burden in immunized mice [979899100]. Multi-epitope-based vaccines, carry the potential B and T cell epitopes simultaneously that relevant with the targeted antigens or helper epitopes. Also, these vaccines with reduction of undesirable factors lead to improve the highly specific responses and better protection [9598].
Go to :
Conclusion
T. gondii can infect a wide spectrum of warm-blooded vertebrate species. Toxoplasmosis predominantly is asymptomatic in normal persons, while may display severe complications with life-threatening outcomes in immunocompromised individuals. Since current drugs have no effect on the encysted parasites, thus, discover of an appropriate vaccine has high priority and urgently required to prevent and control toxoplasmosis, because of the heavy burden of chronic infection worldwide. The use of DNA vaccines expressing CDPK1, CDPK3, and CDPK5, prolonged the survival time as well as reduced the percentage of brain tissue cysts (up to 73.5% reduction) in mice. Nevertheless, these experiments failed to show complete protection (Table 2). The use of live-attenuated vectors as vehicles and prime-boost regimens are other powerful strategies in vaccine design studies which should be paid more attention in future investigations. The complex life cycle of T. gondii indicates the presence of a plurality of antigenic epitopes. As confirmed earlier, vaccination with stage-specific antigens often lead to stage-limited protection. Hence, those antigens that are expressed in various stages of Toxoplasma life cycles, confers more protective immunity against parasite infection [95101]. Since the immunogenicity of the predicted sequences by bioinformatics softwares should be confirmed in suitable animal models, thus, design of investigations with both in vivo and in silico methods is highly recommended to assess the potency of vaccine candidates for the future. Collectively, valuable finding has been gained that gives promising perspectives. Notably, various limitations may affect the outcomes of vaccine studies, including inadequate evaluation criteria, unsuitable vaccination protocol, the strain of T. gondii, the vaccine constructs, the delivery route, dosage of inoculum, various mouse models, and so on. The future surveys must be covered all above mentioned aspects to minimize the faults. Besides, optimized vaccination protocol and use of different types of delivery systems, genetic and/or nongenetic adjuvants undoubtedly would influence the results.
Go to :
 XML Download
XML Download