

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
After Paul Langerhans firstly described dendritic cells (DCs) as nerve cells in 1868, Steinman and Cohn firstly discovered DCs in 1973 as a large stellate or tree-like cell with dendritic morphology of prominent cytoplasmic veils and protrusions [1]. DCs are distinguished from other cells by their unique features, so called dendrites delivering the original name [2]. DCs are broadly expressed in the body but sparsely [3]. They are extremely proficient antigen presenting cells (APCs) that are fundamental to the adaptive immune responses [345]. DCs are not only critical for the induction of primary immune responses, but also important for the induction of immunological tolerance, as well as for the regulation of the type of T cell-mediated immune response [3678].
DCs are derived from hematopoietic bone marrow progenitors and initially transform into immature DCs [910]. These cells are characterized by high endocytic activity and low T-cell activation potential [7]. After DCs contact with an antigen, they become mature DCs and migrate to the adjacent lymph node (LN) [7]. Immature DCs can capture foreign antigens and degrade them into peptide level, and undergo the process which presents the antigen to cell surface by MHC molecules [11]. After antigen capture during inflammation or infection, immature DCs undergo a complex maturation process through Toll-like receptors (TLRs) or members of tumor necrosis factor receptor family. DCs upregulate cell surface costimulatory molecules in T-cell activation such as CD80, CD86, and CD40 highly enhancing their ability to activate T cells [12]. DCs also upregulate a chemotactic receptor such as C-C chemokine receptor type 7 (CCR7) that induces the DCs into the blood stream to the peripheral lymphoid organ or via the lymphatic vessel to lymph node. Finally, DCs activate T helper (Th) cells and natural killer (NK) cells as well as B cells by presenting the antigens together with costimulatory signals [13].
The tumor microenvironment promotes immune tolerance and innovative approaches are required to stimulate immunological antitumor activity or modulate favorable immune system. DCs are the most authoritative candidates which can be helpful to promote immune responses in tumor microenvironment. DCs promote tumor tolerance and modulate the imbalance of Th cells responses. Mature DCs enable to activate T helper 1 (Th1) cells and antigen specific CD8+ cytotoxic T lymphocytes (CTL) under the tumor microenvironment. In this review, we will discuss fundamental features of DCs and the roles of DC-based vaccine as promising cancer therapy.
The Features of DCs
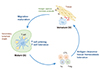
Migration of DCs is an indispensable process before DCs initiate their immune responses (Fig. 1) [14]. Most DCs circulate in the body as an “immature” state. Even though immature DCs lack important functions to induce a strong T-cell response, they are ideally controlled and well equipped to capture antigens and microbes [315]. Once they have acquired and processed the foreign antigens, DCs move to the T-cell areas in peripheral lymphoid organs such as LNs and the spleen, and undergo maturation and stimulate immune responses [16]. Circulating DCs and their precursors exit the blood via tissue-specific recruitment signals such as chemokines that derive from sites of inflammation [17]. DCs are located at surfaces where antigens gain access to the body. DCs are also positioned in distinct incoming channels, called lymphatic vessels, which allow cells to move from peripheral tissues to lymphoid organs [161819]. DCs can encounter immune lymphocytes, selecting those cells that specifically recognize the antigens being carried by the DCs [20]. The lymphocytes begin to grow vigorously and they start to produce materials that will serve to eliminate infections and other sources of antigens [20].
Conventional/Classical DCs
Conventional or classical dendritic cells (cDCs) function as efficient APCs and can induce immune activation or promote tolerance. cDCs have a characteristic morphology defined by long dendrite extensions, and high levels of CD11c and MHC class II expression [2122]. Human cDCs are found in both lymphoid and peripheral tissues [21]. Although there appears to be functional homology between human and mouse cDCs, these cells express their own unique markers, respectively. Two subsets of human cDCs have been characterized that are Lin− (CD3−, CD14−, CD19−, CD20−, and CD56−) and either CD1c/BDCA-1+ or CD141/BDCA-3+ [2324]. CD1c/BDCA-1+ DCs promote a Th1 immune response and act in a tolerogenic manner in response to Escherichia coli [23]. CD141/BDCA-3+ DCs also cross-present extracellular antigens to CD8+ T cells, promote CTL lymphocytes activation, and induce a Th1 immune response [23].
Mouse resident cDCs are found in the central and peripheral lymphoid organs including thymus, spleen, LNs, and Payer's patches. Mouse cDCs comprise two subsets as CD8− cDCs and CD8+ cDCs [2526]. CD8− cDCs can increase MHC class II-mediated presentation of exogenous antigen. Like human CD141/BDCA-3+ cDCs, CD8+ cDCs are able to induce antigen cross-presentation to CTL lymphocytes [25]. Mouse CD8α+ cDCs show their ability to cross-present extracellular antigens to CTLs [27]. They act to maintain tolerance in the steady state and produce IL-12 and interferon (IFN)-γ upon activation [28]. CD8α− cDCs are efficient activators of CD4+ T cells. CD4+CD8α− cDCs comprise a significant proportion of cDCs localized to the spleen, while CD4−CD8α− cDCs account for a significant proportion of cDCs present in mucosal-associated lymphoid tissues [29]. In addition to the lymphoid-resident cDCs, two subsets of mouse migratory cDCs have also been identified that are either Integrin αE/CD103+ or Integrin αM/CD11b+ [30]. Integrin αE/CD103+ cDCs enable to cross-present antigens to CTL cells [31]. They mediate immune tolerance or induce Th2 immune responses. Integrin αM/CD11b+s are found in most tissues including the lung, intestine, and skin [30].
Plasmacytoid DCs
Plasmacytoid DCs (pDCs) are rare subset of DCs that is specialized in type I interferon production [32333435]. pDCs have round lymphocytic morphology and express low level of MHC class II and costimulatory molecules [3637]. pDCs develop in the bone marrow from Flt3+ c-Kitlow progenitors including lymphoid progenitors and common DC progenitors [38]. pDCs are low or negative for CD11c in mouse or human, respectively, but positive for the B-cell marker B220/CD45RA. Particularly, steady-state pDCs are similar to the features of lymphocytes but are different from those of cDCs. Human pDCs express the surface markers blood dendritic cell antigen-2 (BDCA-2; CD303) and immunoglobulin-like transcription-7 [39]. Mouse pDCs express Siglec-H and BST-2/Tetherin [40]. Human IL-3Rα (CD123), BDCA-4 and murine Ly6C, Ly49Q are also useful markers [36]. As components of the innate immune system, pDCs express intracellular TLR7 and TLR9 that detect ssRNA and CpG DNA motifs, respectively [4142]. Upon stimulation and subsequent activation, pDCs produce large amounts of type I interferon (mainly IFN-α and IFN-β), which are pleiotropic anti-viral compounds facilitating various effects [34].
Inflammatory DCs/Monocyte-derived DCs
Monocyte-derived DCs (moDCs) are newly discovered a subset of DCs, which shows common features with classical DCs [43]. moDCs have essential roles in defense mechanisms that induce of both adaptive and innate immune responses [44]. In contrary to cDCs, moDCs are differentiated from Ly6Chigh monocyte progenitors only during inflammatory reactions [45]. However, they share their common features with cDCs; morphology, migration property, priming of T cells, and expression surface markers such as CD11c, MHC II, CD40, CD80, and CD86 [43].
General Properties of DCs
Antigen uptake
DCs are professional antigen processing cells [546]. Immature DCs have several features that allow them to capture antigen. They have a variety of receptors to perform the uptake of antigens, and they are specialized to convert these antigens into MHC-peptide complexes that can be recognized by lymphocytes [4647]. Immature DCs firstly take up antigens by phagocytosis [48]. Next, they form large pinocytic vesicles via a process called macropinocytosis or interaction with a variety of cell surface receptors [49]. Finally, they express receptors that mediate adsorptive endocytosis [4850]. DCs express a variety of receptors that include members of pattern recognition receptors family (such as TLRs, C-type lectin receptors, intracytoplasmic nucleotide oligomerization domain-like receptors), Fc receptors (FcR), complement receptors, mannose receptors and receptors involved in uptake of apoptotic bodies such as phosphatidylserine receptor [515253]. The most prevalent antigen receptors expressed by DCs include members of the C-type lectin family [5455]. For example, DEC-205, a type I C-type lectin containing multiple calcium-dependent binding domains and a unique cytoplasmic tail, may function in directing captured antigens to specialized antigen-processing compartments within DCs [56].
Antigen processing and presentation
Antigen processing by DCs is primarily through two major pathways such as exogenous or endogenous pathway [5]. For exogenous pathway, the captured antigens undergo the endocytic pathway and proteases initiate the degradation of antigens. DCs degrade antigens within a MHC class II-rich endosomal compartment (MIIC) and preserve sufficient peptide structure to be expressed on their cell surface bound to MHC class II, which are produced in endoplasmic reticulum (ER) [5758]. During this process, MIICs change to non-lysosomal vesicles and release the peptide bound MHC complexes on the surface of cells. For endogenous pathway, the phagocytosis and receptor-mediated endocytosis enable antigen uptake, the restricted proteolysis, and the active transport into the cytosol [59]. The cytosolic antigens are additionally degraded via the proteasome. Antigens enter the ER utilizing transporter associated with antigen presenting, and are bound to newly generated MHC class I molecules. MHC class I-peptide is consequently transported by vesicular transport to the surface of cells.
In addition to two antigen-processing pathways, DCs have a specialized antigen-processing process called cross-presentation [60616263]. Antigen cross-presentation describes the process through which DCs obtain exogenous antigens on MHC class I. The antigen is hydrolyzed into oligopeptides after transferring into the cytosol. Then, the antigens are transported to MHC class I molecules in phagosomes or ER [63]. Alternatively, the cleaved antigens by endosomal proteases such as cathepsin S are processed by MHC class I in the endocytic compartment.
Linking of Innate and adaptive immunity
DCs link innate and adaptive immunity by receiving danger signals that render them capable of maturing and inducing productive immunity [764]. DCs also respond to danger signals deriving from foreign substances mentioned to as pathogen-associated molecular patterns that let DCs undergo maturation. TLRs are the most important receptors that recognize microbial products and communicate the information to initiate adaptive immunity. Once primed, the DCs migrate to secondary lymphoid organs, and then present antigens to naïve CD4+ T cells and CTLs. Moreover, DCs release cytokines that further modulate the immune response.
Activation of immune cells
Antigen presentation by DCs is of crucial for the initiation of primary immune responses, due to their unique role in capturing, processing, and transporting antigens [3]. Following activation by antigen encounter, DCs migrate from tissues into LNs. During this process, DC upregulates MHC class I and II as well as costimulatory molecules such as B7.1 and B7.2. This is important for the activation of naïve T cells because they require both signaling the antigen-specific T-cell receptor (TCR) interaction and the costimulatory B7/CD28 mediated second signal. In addition, adhesion molecules such as intercellular adhesion molecule-1 and lymphocyte function-associated antigen 1 are also highly expressed by DCs and induce prolonged cell to cell interaction. They allow naïve T cells differentiate into effector cells. Productive activation of naïve T cells by DCs results in clonal expansion and the effector and memory T cells differentiation [65].
Besides T cell stimulation, DCs are able to stimulate of B cells where happens in LN and germinal centers. Antigens are captured by FcRγIIB can be reserved in intracellular vesicles and presented to B cells [66]. DCs also modulate the functions of NK cells and CTL cells and influence the immune response [67].
Immune tolerance
DCs also induce immune tolerance in both central and peripheral lymphoid organs. They are involved in the important regulatory mechanisms that are clonal selection in thymus, TCR/B-cell receptor editing and regulatory T cells (Tregs) generation [76869]. DCs allow the immune system to tolerate harmless antigens that are originated from own body's tissues, cells and proteins [70]. This is essential to inhibit the body from self-immune response. During the T-cell development in the thymus, DCs contribute in eliminating those cells bearing self-reactive antigens via a mechanism known as central tolerance. DCs also regulate the mechanisms of peripheral tolerance that represent T-cell death, T-cell anergy, and active suppression by Treg [6869]. Proficient immune responses occur when DCs present optimal levels of MHC-peptide complexes with costimulatory molecules. In the event, T cells recognize only low levels of MHC-peptide and have a low affinity for their cognate ligand, or receive no costimulation from DCs, they become anergic or undergo apoptosis [7172].
DC-Based Vaccines as Cancer Therapy
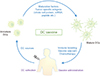
DCs are considered as a leading light of the immune system that connects between innate immunity and adaptive immunity. As mentioned previously, once DC meet the antigen, the antigen is processed and degraded into small peptide to express on the cell surface. These unique functions allow DC-based vaccines to introduce the potent immunotherapeutics for the patients suffering from serious disease such as cancer (Fig. 2) [7374]. DC-based vaccines can be generally categorized by the matured autologous monocytes in vitro or ex vivo, and an antigen-stimulated DCs just before injection [757677]. DC-based immunotherapy is theoretically safe and can promote antitumor immune responses and prolonged survival of patients [78]. DC-based vaccines aims to initiate the immune responses by the stimulation of T cells that can destroy cancer cells and the induction of the memory cells to prevent cancer recurrence [77]. Currently, there exist two types of DC-based vaccines. The one is ex vivo antigen-loaded DC-based vaccines and the other is in vivo DC-targeted vaccines [79]. For in vivo DC-targeted vaccine, it can be divided again according to the types of reactive targets. The first target is ligand such as TLR agonists, and the other is antibody such as DEC205, DC-SIGN, CD11c, and FcγR [80]. In addition, new target is being focused on the delivery system using nanoparticles even though this system is still need to be improved [8182].
DCs for vaccine therapy can be generated from monocytes or CD34+ progenitor cells by the stimulation of cytokines and granulocyte-colony stimulating factor culture medium ex vivo [83]. moDCs are more advantageous than CD34+ progenitor derived DCs because they are possible to be fully differentiated and homogeneous [84]. moDCs are frequently applied for clinical application or trial, and they induce efficient immune responses by tumor-derived antigens.
In tumor microenvironment, DCs can take up damaged tumor cells and then can respond tumor-specific immune responses according to the inflammatory signals. The population of myeloid-derived suppressor cells and Treg keeps high levels at the steady state which means that tumor cells are suppressive [85]. However, the maturation of DCs is inhibited by the specific factors such as vascular endothelial growth factor, interleukin 10 resulting in T-cell anergy and tumor progression [86]. Under tumor microenvironment, the maturation of DCs and the production of mature DCs are decreased and impair tumor suppressive immune response [87]. Also, cancerous cells prevent the efficient antigen presentation and recognition of cytotoxic effects by the conversion of monocytes into macrophages instead of DCs [88].
Recently, DC-based immunotherapy has been focused as a therapeutic approach for cancer treatment. The ability of DCs against tumor has been proven and DCs are produced from monocytes with granulocyte-macrophage colony-stimulating factor/interleukin 4 in vitro, and tumor-derived antigens or tumor mRNA-loaded DCs were directly injected into patients as cancer vaccine resulting the antitumor immune response or tumor killing ability of DCs [8789]. DC-based immunotherapy can be useful for the treatment of unique diseases such as cancer basically depends on functional and immunobiology of DCs.
Conclusion
During more than four decades, the importance of DCs finally has been accepted as indispensable immune cells that connect between innate and adaptive immunity. The prospective roles of DCs as an immunotherapeutic tool are having been effectively resumed by verifying the limitation of cellular and molecular mechanisms of DCs. DC based therapy is supposed to influence to the leader of cancer immunotherapy.
 XML Download
XML Download