

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Influenza A viruses (IAVs) are among the most unpredictable pathogen that threatens human and animal health by continuous evolution through antigenic drift and shift. While the natural reservoirs of IAVs have been known as aquatic wild birds (order Anseriformes or Charadriiformes), the viruses can spill over to susceptible host such as terrestrial birds and mammals including humans [12]. Through complex interactions between viral, host, and environmental factors that determine susceptibility and transmissibility of IAV [34], the cross-species infection can be either transient or endemic or epidemic or even occasionally pandemic [5678910].
Multiple spill-over events and viral adaptation process have resulted in the establishment of stable lineages of IAVs per susceptible host: human (H1N1, H1N2, H2N2, and H3N2), swine (H1N1, H1N2, and H3N2), equine (H3N8 and H7N7), and domestic poultry (many subtypes, H9N2, H6N1, and etc.). Most endemic IAV infections are clinically asymptomatic or cause mild respiratory signs; those endemic strains have been known as seasonal flu viruses in human [1112] and as low-pathogenic avian influenza viruses (LPAIVs) in poultry [13]. However, continuous emergence of novel IAV reassortants from non-human hosts such as pigs and poultry, which is possibly susceptible to human population but have not been exposed before poses a potential pandemic threat to public health [214]. Furthermore, in domestic poultry, highly-pathogenic avian influenza virus (HPAIV) can arise from H5 and H7 LPAIV via acquisition of multibasic amino acid cleavage sites of hemagglutinin (HA) molecule [1516] and intercontinental spreads by migratory birds have made HPAIV as a global panzootic disease [1718].
Considering the substantial threats of IAVs to human and animal health, the viruses are monitored and controlled through a global collaboration of world organizations such as World Organisation for Animal Health (OIE), Food and Agriculture Organization of the United Nations (FAO), and World Health Organization in the concept of “One Health” [41920]. OFFLU, the OIE/FAO network of expertise on animal influenza, was developed and the vision of the consortium is as follows. The animal health community will provide early recognition and characterization of emerging influenza viral strains in animal populations, and effective management of known infections, thereby better managing the risk to human health and promoting global food security, animal health and welfare, and other community benefits derived from domestic animals and wildlife.
Vaccination is considered the most effective tool for the control and prevention of IAV infections in both humans and animals. However, especially for livestock animals in industrial settings (e.g., rapid turnover and high-density stocking), careful applications of IAV vaccines are required with a high level of biosecurity control and periodic surveillance for the vaccines not to work as driving forces that accelerate the viral evolution. While inactivated vaccines have been used conventionally worldwide in human and animal medicine, new generation IAV vaccines adopting recent technologies have been being developed and commercialized. Here, we review challenges of influenza virus that humans and animals face today highlighting specific concerns for each host species (human, domestic poultry, and swine) and discuss influenza vaccines in their usages, application strategies, and developments.
Go to : 
Seasonal, Pandemic, and Zoonotic Influenza Viruses in Humans
Influenza infection in humans can be classified into seasonal, pandemic, and zoonotic form. Seasonal flu is caused by influenza A, B, and C viruses and accounts for about 3 to 5 million cases of severe illness, and about 250,000 to 500,000 of deaths annually worldwide [21]. It has been estimated that the total annual economic burdens of seasonal influenza epidemics amounted to $87.1 billion [22]. Of three types of influenza, type A is known most virulent. Genetic subtypes of currently circulating seasonal IAV strains are H1N1 and H3N2 [23] and the epidemics have been dominated by H3N2 since 1968 when the “Hong Kong flu” occurred [24].
Influenza pandemic occurs when people are immunologically naïve to a newly emerged IAV and the virus exhibits sustainable transmissibility between humans. A total of four influenza pandemics have occurred since the 20th century: Spanish flu pandemic (H1N1, 1918–1920), Asian flu pandemic (H2N2, 1957–1958), Hong Kong flu pandemic (H3N2, 1968–1969), and swine flu pandemic (H1N1pdm09, 2009–2010). The Spanish flu is among the most devastating pandemic, having killed at least 50 million people. Subsequent pandemics have resulted in many fewer fatalities: the Asian flu (1.5 million), the Hong Kong flu (1 million), the swine flu (>18,500) [2]. The causative viral strain for each epidemic has been emerged from genomic reassorment within avian host (Asian flu and Hong Kong flu) or porcine host (swine flu). In case of Spanish flu, it is still controversial whether the virus had been a reassortant generated in mammals or a directly adapted avian influenza that crossed the species barrier from birds to human [2526272829]. The several descendants of pandemic influenza virus strains have established a novel lineage in humans and become a regularly circulating seasonal flu virus (e.g., H1N1, H3N2, and H1N1pdm09) [30].
Zoonotic influenza viruses are similar to pandemic influenza viruses in the immune status of human populations against the newly emerged viruses, except that the infections by those viruses end as sporadic individual events with very limited transmission to others. Although avian or pig influenza viruses do not infect humans in general, sporadic human infection can be made via direct contact with infected animals or contaminated environments. Until now, human infections by avian influenza (H5N1, H5N6, H7N7, H7N9, H9N2, and H10N8) [31] and swine influenza (variant H1N1, H1N2, and H3N2) have been reported [3233]. The disease severity in human varies according to viral strain; high lethality has been recorded in H7N9 (612 deaths among 1,565 infected patients since 2008), H5N1 (454 deaths among 860 infected patients since 2003), and H5N6 (7 deaths among 17 infected patients since 2016) [34]. H5N1 cases have been reported in 16 different countries distributed across Southeast Asia to Eastern Europe with Egypt and Indonesia reporting most of the infections. On the other hand, most reports of human infections by H7N9 and H5N6 have been limited in China. Despite the mild clinical illness and rapid recover in infected humans, H9N2 seems to have immense importance in terms of epidemiology of zoonotic influenza because the virus has been recognized as a potential enabler virus. The hypothesis was originated from the finding that H9N2 was a common denominator contributing to the genesis of H5N1, H5N6, H7N7, and H10N8, zoonotic viral strains as mentioned above, by providing whole sets of internal gene (PB2, PB1, PA, NP, M, and NS) [231]. It has been reported that human infections by variant swine influenza viruses (SIVs) tend to result in mild clinical illness [34].
Go to :
Avian Influenza, A Global Devastating Disease to Poultry Industry
As per pathotypes in terrestrial birds, avian influenza viruses can be classified into LPAIV and HPAIV. Aquatic wild bird has been known as a reservoir for LPAIV [35] and is much likely to show clinical resistance to HPAIV as well as LPAIV than is domestic poultry, even considering that H5N1 strain represents high morbidity and mortality in wild species [3637]. In domestic poultry, HPAIVs cause severe disease, with fatality as high as 100%. In recent decade, global highly-pathogenic avian influenza (HPAI) outbreaks have been dominated by subtype H5 IAV strains (H5N1, H5N2, H5N6, and H5N8), all of which was derived from A/goose/Guangdong/1/96 (H5N1) [38]. Disease outbreaks of the A/goose/Guangdong/1/96 lineage viruses had been confined to the Asian continent before 2005. However, the viral spreads out of Asia were identified in Africa and Europe in 2005 (clade 2.2) and even in Americas in late 2014 (clade 2.3.4.4), which have been suspected of being spread by migratory birds [3839].
The disease outbreaks have severely affected nation's poultry industry by causing tremendous economic losses including direct and indirect costs. Direct costs can be derived from (1) production losses by the disease itself and a series of actions to stabilize the onset of outbreaks (e.g., stamping-out, culling, and vaccination) and (2) control costs during and after disease outbreaks. The costs vary between countries because it is determined by the contribution of poultry industry to the national gross domestic product (GDP) and governmental control policies such as stamping-out and/or vaccination. Indirect costs include ripple effects, spill-over effects, and potential costs occurring with zoonotic events in related business and general public. Detailed definitions and examples for each cost are summarized in Table 1[40]. It is generally considered that the indirect impact is much greater than the direct impact because trade, tourism, and services that can be affected by public fear of zoonotic potential and international regulations have a large contribution to the national GDP.
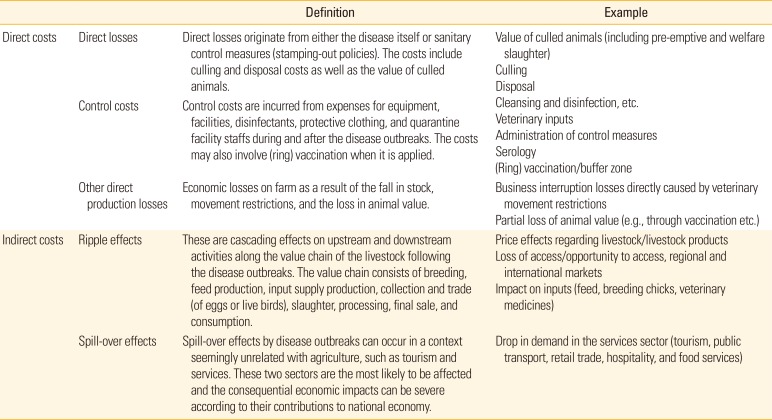
Table 1
Definitions and examples of disease outbreak costs
Modified from the World Organisation for Animal Health (2007), with permission of World Organisation for Animal Health [40].
![]()
Over 150 million poultry were killed in Asia during 2003–2004 HPAI H5N1 outbreaks and as a result, the total GDP losses were estimated at US $10 billion. Of countries affected by severe economic losses, Thailand suffered the largest economic losses recording total estimates of about US $1.2 billion. While the direct losses of Thailand (29 million birds, 14.5% of the poultry population) were the second-highest after Vietnam (44 million birds, 17.5% of the poultry population), trade losses were so severe that the largest total costs were recorded in Thailand, which was the inevitable result given that the country had been the world's fifth largest poultry exporter in the world. In HPAI outbreaks in United States during 2014–2015, economy-wide total loss estimates were $3.3 billion that includes $1.6 billion of direct costs from destroyed turkeys and egg production and partial indirect costs by negative knock-on effects on wholesale, retail, and food service sectors [41]. In Korea, the economic losses associated with HPAI outbreaks were largest in 2016–2017 since the first HPAI outbreak in 2003, with expected estimates of at least $435 million and maximum of $1.3 billion [42].
A survey of OIE member countries found that more than a quarter of the major disease outbreaks that occurred worldwide from 2000 to around 2016 (358 outbreaks) were occupied by avian influenza and those flu outbreaks were the most frequently related with trade issues [43]. When HPAI outbreaks occur in an exporting country, sudden unexpected trade bans are imposed by importing countries, which can result in severe economic losses to the exporter. To minimize the losses as well as prevent the viral spreads via trade commodities, despite the controversial issues about the applicability in other countries, the United States and European Union (EU) countries have been implementing ‘regionalization’ that reduces restrictions from whole country to smaller zones/regions. The regionalization should be based on comprehensive surveillance and risk assessments as recommended by the OIE Terrestrial Animal Health [13]. During HPAI outbreaks in United States during 2014–2017, successful applications of regionalization for the trade of poultry products have been reported; especially, at a single outbreak of H7N8 HPAI in the state of Indiana, many trading partners with United States did not impose restrictions as such large of a zone or wide of a range of products as they did during 2014–2015 HPAI outbreaks [44].
Go to :
Pigs as Genetic Reservoirs for IAVs
While SIVs cause acute respiratory signs in pigs, the economic impacts of SIVs on swine industry as itself are not as high as those of HPAIVs on poultry industry because the infected pigs are frequently subclinical and characterized by low mortality (<1%) with rapid recovery in uncomplicated conditions [45]. However, substantial economic losses can occur in case of viral transmission of pandemic influenza from pigs to humans. Although the impacts of the 2009 H1N1 pandemic on global economy was estimated to be less than 1 percent of GDP, the costs for possible pandemics in the future could increase [46].
Cell surface receptors for both human (α2,3-linked sialic acids) and avian influenza viruses (α2,6-linked sialic acids) have been identified on swine respiratory epithelial cells, suggesting that pigs can be responsible for the emergence of pandemic influenza by serving as “mixing vessels” [47]. For the reason, swine influenza has attracted much more attentions in terms of public health rather than economic aspects. Recently, contrary to the prevailing belief, limited expression of α2,3-linked sialic acids on upper respiratory tracts of pigs has been identified in several studies [4849]; accordingly, the importance of pigs as mixing vessels was somewhat diminished. However, the resemblance of receptor distribution (α2,3- and α2,6-linked sialic acids) between human and pig still poses substantial roles of pigs in emerging pandemic threats. Actually, H1N1pdm09 has a genomic constellation of swine origin; six segments (PB2, PB1, PA, HA, NP, and NS) from triple reassortants of North American swine lineage and the other two segments (M and NA) from a Eurasian swine lineage [50]. Moreover, following reverse zoonosis (spread from human to animal) of H1N1pdm09 to swine populations, as a consequence of reassortment of H1N1pdm09 with endemic swine H3N2, variant H3N2 (H3N2v) strains containing H1N1pdm09 internal genes (particularly, PA, NP, and M gene) have emerged and resulted in continuous human outbreaks in the United States since the first report in July 2011 [3351]. The highest number of human outbreaks from July 2011 to December 2017 in the United States has been reported for H3N2v (H3N2v, n=434; H1N1v, n=21; H1N2v, n=13) and the case counts are as follows: 12 (2011), 309 (2012), 19 (2013), 3 (2014), 3 (2015), 18 (2016), and 62 (2017) (https://www.cdc.gov/flu/swineflu/h3n2v-case-count.htm). Fortunately, it has been known that H3N2v strains reported until now have limited transmissibility between humans.
The pig population performs as genetic reservoirs for IAVs that had been prevalent in human population and manufacturers of new IAV strains. It has been reported that even though human seasonal H1N1 strains were replaced by H1N1pdm09, H1s of previous seasonal flu viruses still circulates among swine populations of North America, Europe, Asia, and South America [952]. Similarly, in China, the circulation of the parental strain of seasonal influenza virus H3N2 was confirmed in pigs long after the strain had been replaced by other strains in humans [53]. The genetic information of human seasonal influenza viruses preserved in swine population by region or country is summarized in Table 2. The longer human-origin IAVs circulate within pigs without exposure to humans, the more likely that immunological gaps will be made between older and younger generations, which makes that young people become immunologically vulnerable to previous season flu viruses, which they may encounter during their lifetime.
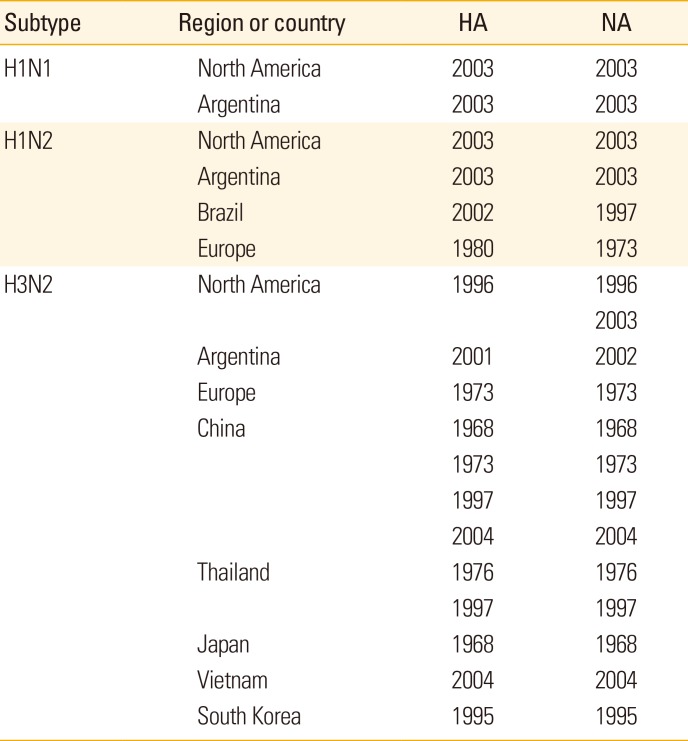
Table 2
Influenza A viruses (IAVs) in swine by region originated from human seasonal IAVs with isolation year of the most closely related genetic ancestora)
Modified from Vincent et al. Zoonoses Public Health 2014;61:4-17, with permission of John Wiley & Sons [9].
HA, hemagglutinin; NA, neuraminidase.
a)The information of H1N1pdm09 transmitted from human to pig was excluded.
![]()
Go to :
Vaccines Against IAV and Their World Usage in Animals
As the global human population continues to grow with the demand for animal proteins, the population density of domestic stocks such as chicken, ducks, and pigs has been increasing in many parts of the world including Asia and the United States [54]. Given the economic burdens for IAV outbreaks and the complexity of host-virus interplay, effective and thorough control strategies in animal sectors are crucial and necessary.
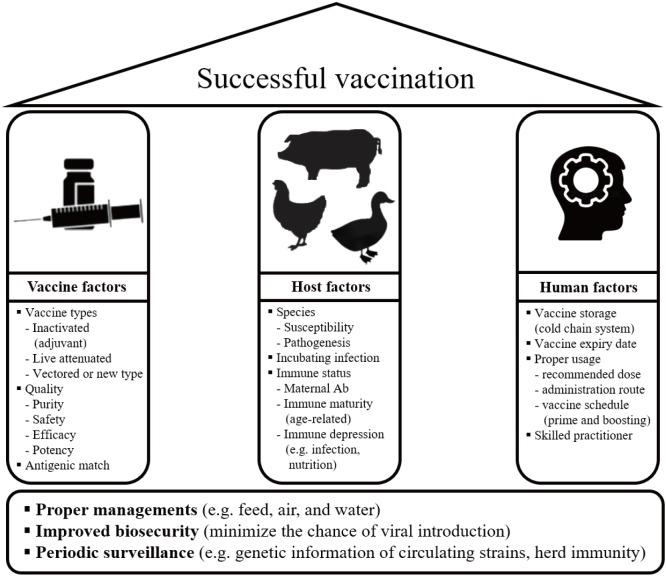
Vaccination is regarded as a cost-effective and efficient tool for the control of IAVs in animals. However, as a rule of thumb, a high level of biosecurity that includes all the related activities to control and prevent viral spreads such as changes of rearing style/facilities, disinfection, movement confinement must be accompanied with vaccination strategy based on sufficient surveillance data (Fig. 1)[5556]. Biosecurity measures also include interventions to reduce likelihood of IAV transmissions between humans and animals at animal-human interface such as abattoir and wild poultry markets in Asia [57] and additionally, pig agricultural fairs in the United States [58].
Vaccine usages in poultry industry
Currently, vaccination of domestic poultry (chicken and turkey) is commonly implemented against HPAIV, H5/H7 LPAIV, and H9N2 LPAIV worldwide. Historically, emergency vaccination against HPAIV to control epizootics has been used in Mexico (H5N1, 1995–1995), Pakistan (H7N3, 1995–2004), Asia/ Africa/Europe (H5N1, 1996–continuing), and North Korea (H7N7, 2005) as an adjunction to stamping out programs. Meanwhile, China, Egypt, Vietnam, Indonesia, and Hong Kong have adopted routine vaccination of all poultry within a country against H5N1 HPAIV since 2002, which accounts for 99% of world total vaccine usage for HPAIV control from 2002 to 2010. The percent of vaccine usages and average coverage rate by countries as follows: (1) vaccine usage: China (90.99), Egypt (4.65), Hong Kong (0.08), Indonesia (2.32), and Vietnam (1.43) and (2) vaccine coverage rate: China (47.1), Egypt (69.9), Hong Kong (86.2), Indonesia (14.0), and Vietnam (52.3) [5960].
The usage of vaccines containing H5 or H7 antigen against LPAIV was identified in six countries (Mexico, Italy, Guatemala, El Salvador, Portugal, and the United States) but the total amounts of doses (10.1 billion) are much smaller than those used for HPAI control (>113 billion). Due to relatively strong field impacts (decline in egg production and moderate to high mortality) among LPAIVs, H9N2 also has been controlled via vaccination in the Middle East and Asia including South Korea, Pakistan, and Israel [5960616263].
Vaccine usages in swine industry
Vaccine usage to control swine influenza varies by countries; some countries use vaccination strategies, while others do not. For examples, SIV vaccination has been conducted extensively in the United States. On the contrary, in Korea, vaccines for SIV control have been rarely used despite the presence of a commercially available vaccine. Due to genetic diversity of circulating SIV strains, most commercial vaccines consist of multiple strains of subtype H1N1, H1N2, and H3N2. However, the fast evolution of field viruses could surpass the updates of commercial vaccines. Accordingly, combined use of herd-specific autogenous vaccine with other commercialized vaccines is implemented in some countries; about 20% of pig farms in the United States used the autogenous vaccines in 2006 [646566]. However, compared to avian influenza viruses, it seems that vaccines against SIVs have not been utilized actively by swine veterinarians in many countries (at least in Asia) because deleterious effects on swine health by other major pathogens such as porcine reproductive syndrome virus and porcine circovirus are so severe that SIV has not been noticed as a disease that needs control.
Go to :
Considerations to be Successful Vaccination
For successful IAV vaccination, several factors related with vaccine, host, and human should be considered and harmonized (Fig. 1)[6768]. Most of currently licensed vaccines worldwide are the classic type of vaccine containing inactivated whole virus antigen formulated into mineral oil emulsions. Although several new effective generation vaccines have been developed [69], time will be required to resolve safety, efficacy, and cost issues in the field application and finally, for the vaccines to be licensed, which highlights the necessities of the effective and proper use of the classical inactivated vaccines.
High quality of vaccine and proper delivery to appropriate hosts are prerequisites for successful preventive vaccination, which can be assessed by antigen-specific response in vaccinated animals [70]. The surface glycoprotein HA is the major target of neutralizing antibodies (nAbs) against IAVs and the antibody quantities can be measured using hemagglutination inhibition (HI) assay utilizing the trait of the nAb that inhibits red blood cell agglutination. The major determinant of vaccine for success is antigenic match between circulating virus and vaccine strains. Vaccine strain homologous with field strains may confer the best protection against field strains of virus. For heterologous field strains, HI assay with host sera specific to vaccine antigens gives guidance to vaccine strain selection [71]. Moreover, it is anticipated that recent computational technologies such as antigenic cartography and antibody landscape modelling can provide fast, cost-effective and accurate analyses of candidate vaccine strains [56].
Inaccurate antigenic matching between vaccine strains and field strains leads to lower protective immunity and facilitates the selection of more resistant progenies or phenotypes to vaccine-induced immune conditions rather than elimination [72]. Consequently, the evolution of viral population into vaccine-escaping directions such as emergence of immune escaping mutants and change of dominant viral genotype occurs. Those phenomena have been observed both in avian virus [737475] and SIVs [7677] as well as in other viruses of industrial animals [78798081]. Incomplete vaccine coverage commonly observed in the fields also can be a critical contributor accelerating the evolution [82].
Recently, vaccine-associated enhanced respiratory disease (VAERD) has been identified in vaccinated pigs against swine IAVs. When pigs vaccinated with oil-in-water adjuvanted whole inactivated virus were challenged with viruses of same subtype but distinct antigenicity, more severe clinical signs and lesions of inflammation in lungs were observed compared to non-vaccinated pigs [838485]. Multifactorial pathological mechanisms may be involved in VAERD. However, the vaccine-induced IgG antibodies that fails to cross-neutralize mismatched challenge strains seem to mediate enhanced pathogenesis [86].
Along with the good quality of vaccine, also crucial are host factors for successful preventive vaccination. Basically, animal herds to be vaccinated should not be in immunologically compromised conditions. Infections with immunosuppressive pathogens (e.g., infectious bursal disease and reticuloendotheliosis virus in chickens) and poor management practices (nutrition depletion, unhygienic condition, and etc.) can hamper vaccine-induced immune responses [7087]. Maternally derived antibodies (MDA) can also interfere with vaccination. Additionally, it has been experimentally demonstrated that MDA from sows immunized with whole inactivated vaccine (WIV) can make their offspring become susceptible to VARED upon heterologous infection [88], which implies the importance of vaccine strain selection in pigs.
Go to :
Limitations of WIV
Since the WIV stimulates mostly humoral response rather than cell-mediated response [89], sterilizing immunity is unlikely to occur in WIV vaccinated animals even to challenged viruses homologous with the vaccine strain; although clinical manifestations and viral shedding are significantly reduced with pre-immunization, the induced immunity cannot prevent infection [90]. Silent spread of HPAIV H5N1 without clinical signs in vaccinated flocks with WIV, which is the main reason for the reluctance to HPAIV vaccination in many countries and communities, is an example showing disadvantage of non-sterile immunity induced by WIV [9192].
Dynamic changes in IAV epidemiology require corresponding rapid responses including updating of vaccine strains. Currently, most flu vaccines in human and animal medicine are produced in chicken embryos, which means that the vaccine strains need to grow well in chicken eggs. However, in case of HPAIVs, those viruses isolated from nature do not grow well in eggs because of early embryonic death. Therefore, attenuation procedure using reverse genetics is essential with following quality assurance and pathogenicity test in animals, which takes several months (about 22 weeks) to create new vaccine strains [93]. To overcome this and other several issues related with egg-based production systems such as egg supply and inflexibility with modern biotechnologies, cell-based vaccine production using fixed cell lines has been developed [9495]. In 2016, Food and Drug Administration of the United States approved the use of cell-based viruses for the production of human influenza vaccine (Flucelvax) [9697].
Due to non-infectious nature of killed antigen, delivery of WIV must be provided via intramuscular or intradermal routes. Considering high density rearing and rapid turnover rate of livestock, especially pigs and poultry in industrial settings, the individual vaccination is highly laborious and error prone process, leading to incomplete herd immunity and vaccine failures. On the contrary, mass application with reduced vaccination time and labor cost is available using water supply or aerosol spray of infectious viruses such as attenuated live viruses [98] and recombinant Newcastle disease viruses (NDV) [99100101102].
Go to :
Next Generation IAV Vaccines in Avian and Swine Industry
Vectored vaccines
Vectored vaccine has been investigated extensively using various viral vectors: human adenovirus 5 and pseudorabies virus in pigs [103]; fowlpox virus (FPV), NDV, herpesvirus of turkey (HVT), duck enteritis virus and etc. in poultry [104]. Among them, vectored vaccines using FPV, HVT, and NDV have been commercially available in more than one country [104]. Vectored vaccine uses viral vectors to deliver target protein to host's immune system. HA protein is mainly targeted in vaccines against IAV. Because the vectors infect host cells, those safely activate both humoral and cell-mediated immunity. However, if the host have maternal antibody or preexisting immunity to vector, the vectored vaccines fail to immunize host against IAV. For example, because routine vaccination programme against NDV (avian paramyxovirus serotype 1, APMV-1) has been implemented in many countries, most commercial chickens have pre-existing immunity against NDV vectors. As alternative measures, other serotypes of APMV having low cross-reactivity with anti-NDV antibody such as APMV-2, APMV-3, APMV-6, and APMV-10 have been proposed for candidate vectors [105106]. For the same reason, those type of vaccine cannot be administered multiple times; in a prime boost strategy, vectored vaccine should be applied first, followed by other types of vaccine for boosting purpose. Distinct characteristics of vaccines can be exhibited depending on the nature of the viral vectors. For example, FPV vectors are not efficiently interrupted by MDA and can confer protection after single vaccination on 1-day-old chicks [107108]. Like avian influenza viruses, the major infection route of NDV vectors is respiratory tract, which enables the mass vaccination via aerosol spray as mentioned [99109].
RNA vaccines
RNA vaccines can be classified into two types: one is conventional mRNA vaccines and the other is self-amplifying mRNA (SAM) vaccines. The mRNA containing gene of interest that resembles host cell mRNA is the required factor for the conventional mRNA vaccine. The genome of RNA viruses such as alphavirus engineered not to produce viral progenies has been applied in SAM vaccines. Because the gene of target is substituted for viral structural gene which is essential for viral packing, the RNA replicon cannot produce infectious viral particles but expresses high levels of target antigen via self-amplification in host cells [110]. In contrast to DNA, RNA only needs to cross one barrier, plasma membrane, to express target proteins and can be actively taken up by dendritic cells or other cell types via receptor-mediated endocytosis [111], which enables the vaccines to be administered via intramuscular or intradermal route. Delivered RNAs are recognized by innate immunity using pattern recognition receptors, which leads to subsequent activation of cell mediated and humoral immune response [112]. Recently, significant improvements in overall process to make RNA vaccines such as synthesis, stability, purification and delivery of RNA have been achieved [113114115116117118]. Moreover, the simple concept of the vaccine will make it feasible to rapidly reflect the contemporary field IAV strains (estimated as 6-8 weeks from sequence data acquisition to quality control test) [118].
The RNA vaccines using SAM technology for swine influenza have been commercialized in the United States and the number of doses used alongside autogenous vaccines during 2008-2011 accounted for about 50% of total doses of swine influenza vaccines [56]. The SAM vaccines for avian influenza were also recently licensed in the United States [104]. For conventional mRNA vaccines, although there is no commercialized product for IAV yet, its high efficacy has been demonstrated in mouse, ferret, and pig models [118]. The efficacy in poultry species remains to be tested. Most recently, effective seroconversion after the mRNA vaccination against H10N8 and H7N9 has been demonstrated in monkeys and humans [119].
Universal vaccines
The concept of universal vaccine is to cover all subtypes of IAVs. Developing such a vaccine that induce broadly cross-reactive immunity would be beneficial in both human and animal medicines considering dynamic epidemiology and genetic diversity of IAVs. Highly conserved antigenic domain has been the target for the development of universal IAV vaccines. Stalk domain relatively conserved compared to globular head domain in HA and M2 ion channel has been investigated extensively for the promising vaccine candidates [120121122]. NP, PA, and M1 have been also considered to provide heterosubtypic protection. However, as demonstrated by many experimental challenge studies, cross protection conferred by universal IAV vaccine is weak and depends mostly on cytotoxic T lymphocyte responses with the minimal effect of nAbs [123]. Unfortunately, in pigs vaccinated with universal vaccines based on HA stalk domain [86] and extracellular M2 protein [124], exacerbated clinical signs and even deaths were observed, which suggests the careful clinical application of universal vaccine in pigs.
DIVA vaccines
Most types of vaccine except live attenuated vaccine can be compatible with differentiating infected from vaccinated animals (DIVA) strategies, which is essential for the effective disease control in animals. For example, the presence of antibodies against nonstructural protein (mainly NS1), which is absent in animals vaccinated with inactivated IAV can be biomarkers for viral infection [125]. In case of vaccines utilizing particular genes or proteins (mainly HA or NA), only specific immune response against targeted protein is induced and thereby can be used for DIVA. Those vaccines involve nucleic acid (DNA/RNA) vaccines, subunit vaccines, peptide vaccines, virus like particles, and vectored vaccines. Although there exist several live attenuated vaccines available for DIVA (e.g., neuraminidase deleted IAV vaccine), it is not clear that whether those vaccines can be used in the fields [126127].
Go to :
Conclusion
Continuous evolution of IAV is not a risk confined to particular species as described in this review. Human outbreaks by zoonotic and pandemic influenza virus at animal-human borders have been occurring worldwide with regular seasonal flu outbreaks and human influenza viruses have been introduced to swine by reverse zoonosis. Recent HPAI outbreaks have been severely damaging poultry industry and human infections by the HPAIVs have raised concerns for public health. Taking into consideration of dynamic interplay of IAVs between human and animal sectors, vaccine is considered a promising preventive or control tool that could make synergistic effects when combined with proper biosecurity practices.
Different vaccination strategies against HPAI have been used as per country policy. While some countries adopted regular HPAI vaccination, others used the HPAI vaccine only in an emergency and the others stick to stamping-out policy without using vaccines. In South Korea, despite the repetitive HPAI outbreaks with continued economic losses, the government is reluctant to use HPAI vaccines behind the fear of becoming endemic status and concerns about impact on trade. Countries such as China, Indonesia, Egypt, and Vietnam that have implemented vaccination against HPAIV still remain endemic. However, the vaccine in these countries was introduced after HPAI become enzootic. Furthermore, their inadequate vaccine coverage seems to attribute to continued HPAI outbreaks and the emergence of antigenic variant strains. Rather, trade losses by HPAI outbreaks can be minimized through rapid stabilization of outbreaks by the effective use of HPAI vaccines. Those can be accomplished via harmonization between emergency ring vaccination with stamping out policy and high levels of biosecurity accompanied by thorough surveillance. The importance of pigs as genetic reservoirs of human and avian lineages of IAVs has not been taken seriously in many Asian countries including Korea. However, in the One Health concept, it seems that SIVs in Asian pig populations should be monitored and controlled by the supports of governments, field veterinarians, and swine industry.
Over the recent decades, various experimental vaccines against IAV have been developed to enhance the vaccine efficacy and safety. WIV vaccines are the most widely used and safe vaccine but there are concerns about cross protection and low efficient viral elimination. On the contrary, live attenuated vaccines are considered the most efficacious among existing vaccine types due to rather broad coverage of protection and rapid viral clearance but have safety problems related with reversion to virulence. The promising next generation vaccine candidates that have strength both in efficacy and safety seem to be vectored and RNA vaccines. In addition, under the era that requires rapid response against IAV threats, the flexibility of these two vaccines is of great advantage. Every vaccine developed and under development has its pros and cons and there is no absolute vaccine that can be applied to every host and condition effectively at the best performance. In practical aspects, vaccines that are highly compatible with massive immunization methods such as aerosol spray and water supply and DIVA are highly recommended for vaccine usages in animals under industrial settings. Further, complementary vaccine usage (e.g., NDV vectored vaccine for prime and WIV vaccine for boosting) can be an attractive alternative strategy to prevent and control IAVs. We believe that recent developments of vaccine technology will bring us more efficient tools for the prevention and control of IAVs.
Go to :
 XML Download
XML Download