

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Veterinary medicine considers prophylactic vaccines, in combination with strict bio-security, to be the most cost-effective tools for preventing viral infections. In general, veterinary viral vaccines aim to protect susceptible host animals from fatal infectious diseases by inducing a rapid and long-lasting immune response and by preventing the spread of such diseases among populations by reducing viral shedding from infected animals. Such vaccines must be cost-effective, stable, and easy to administer. Live attenuated virus and/or inactivated virus vaccines have been used for decades to prevent viral infectious diseases; however, many vaccines do not satisfy the requirements for an “ideal vaccine” in the field due to limited effectiveness and/or side effects. In addition, no vaccines are available for some infectious diseases.
Advances in recombinant DNA technology have made it possible to design new innovative genetically engineered vaccines with improved safety profiles and greater protective efficacy. These “next generation” vaccines include DNA vaccines, subunit or virus-like particle vaccines, genetically modified marker vaccines, and virus vectored vaccines. Of these, the latter are thought to be a promising tool for developing polyvalent or antigen delivery vaccines that express foreign antigen(s) derived from pathogens of economic importance from a veterinary and human perspective.
Many viruses have been used to develop virus vectored vaccines, which provide effective protective immunity against foreign antigens. The number of vectored vaccines licensed for veterinary and human use has increased over time. Initially, vectored vaccines were based on DNA viruses such as herpesviruses, animal poxviruses, and adenoviruses [1]. Currently, with advances in reverse genetics approaches, many RNA viruses have been adapted for use as vectored vaccines [2]. Such vaccine vectors include both positive-sense RNA viruses (picornaviruses, coronaviruses, and flaviviruses) and negative-sense RNA viruses (paramyxoviruses and orthomyxoviruses) [2]. In particular, Newcastle disease virus (NDV), a paramyxovirus that infects birds, is used as an important vaccine vector for development of bivalent vaccines against pathogens of economic importance to the poultry industry. Numerous studies demonstrate that NDV is a promising vector for delivery of protective antigens derived from pathogens infecting mammals and humans; indeed, NDV vectors are safer and more efficient than conventional whole virus vaccines [3]. This article reviews recent developments in the field of NDV-vectored vaccines and the potential use of such vaccines in veterinary and human medicine.
Newcastle Disease Virus
Virulent NDV, the causative agent of Newcastle disease (ND), results in devastating economic losses to the poultry industry worldwide, particularly in Asia, the Middle East, and Africa [4]. To control ND, poultry flocks are subjected to prophylactic vaccination with live attenuated and/or killed NDV vaccines [5].
Gallinaceous birds (e.g., chickens, quail, and turkeys) are highly susceptible to NDV, although the virus infects most bird species. The clinical severity in chickens varies greatly, depending on the virulence of the NDV isolate. Based on their effects in chickens, isolates of NDV are classified into at least four pathotypes: velogenic (a highly pathogenic form), mesogenic (a moderately pathogenic form), lentogenic (a low pathogenic form), and avirulent (a subclinical form) [6]. Lentogenic (e.g., LaSota and B1) and avirulent (e.g., VG/GA and V4) viruses are widely used as vaccines to prevent ND in poultry.
NDV is an avian paramyxovirus type 1 virus belonging to the genus Avulavirus within the family Paramyxoviridae [7]. The virus contains a nonsegmented, negative-sense, single-stranded RNA genome of 15,186 to 15,198 nucleotides in length, which encodes (3' to 5') six structural proteins: nucleocapsid (N) protein, phosphor (P) protein, matrix (M) protein, fusion (F) protein, hemagglutinin-neuraminidase (HN) protein, and RNA-dependent RNA polymerase (L) protein [8]. The length of the NDV genome follows the “rule of six,” which allows efficient virus replication because the N protein binds effectively to six nucleotides [910].
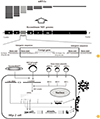
Fig. 1 shows a schematic diagram of NDV morphology. The NDV F and HN glycoproteins are the major structural proteins and comprise a bilayer lipid membrane (envelope) that plays an important role in virus attachment, entry, and release. The M protein resides beneath the envelope and is involved in assembly and budding. The N protein binds to the genomic RNA to form the nucleocapsid (NC) core, and the P and L proteins associate with the NC core to form the ribonucleoprotein (RNP) complex, which is essential for virus replication and transcription [9]. Two nonstructural proteins, V and W, can be produced by RNA editing of the P gene: one (V) or two (W) G residues are inserted at a specific location within the mRNA encoding the P gene [9]. The V protein is an interferon (IFN) antagonist [11].
The life cycle of NDV occurs entirely in the cytoplasm. First, NDV enters the cell cytoplasm via viral attachment and subsequent membrane fusion, which are mediated by the HN and F proteins, respectively. The viral NC or RNP complex is then released into the cytoplasm, whereupon the RNP complex transcribes the viral RNA to produce viral mRNAs, which are then translated into viral proteins. Genome replication occurs when sufficient viral proteins (especially the N protein) are synthesized. The full-length anti-genomic RNA (plus-strand) serves as the template for synthesis of the viral RNA genome (minus-strand). The newly formed genomic RNAs are then encapsidated by the N protein before associating with the polymerase complex. All viral components are then transported to the cell membrane, where they are assembled into progeny virus particles: the progeny acquire an envelope by budding under the direction of the M protein before being released from the cell surface by a process of detachment (elution), which is regulated by the HN protein (which has neuraminidase activity).
In Vitro NDV Recovery System
Because NDV is a negative-sense single-stranded RNA virus, a functional viral RNP complex is required for viral transcription and replication. Thus, the reverse genetics technology used to generate recombinant NDV (rNDV) requires a full-length complementary DNA (cDNA) clone and three helper plasmids encoding the NDV N, P, and L proteins under the transcriptional control of a T7 RNA polymerase promoter [812]. Once simultaneously transfected into permissive cells, the three helper plasmids express the N, P, and L proteins, which then bind to genomic or anti-genomic RNA to form the RNP complex.
In general, reverse genetics approaches used to rescue rNDV are based on one of three different systems: (1) a chicken fibroblast cell (or QM5 cell line) and fowl pox virus-T7 recombinant system [13]; (2) a T7 RNA polymerase expressed BSR T7/5 (baby hamster kidney-21 cell clone) system [14]; or (3) a human epidermoid carcinoma (HEp-2) cell and a host-restricted recombinant vaccinia virus (modified vaccinia virus Ankara [MVA]) system [15]. Fig. 1 depicts the reverse genetics system used in our own laboratory. Briefly, HEp-2 cells are infected with MVA and then cotransfected with a NDV cDNA clone (LaSota backbone) and three helper plasmids (N, P, and L). The recovered viruses are purified on HEp-2 cells in a plaque assay, and clones are selected and passaged several times in specific pathogen free (SPF) embryonated chicken eggs.
Insertion of foreign genes into the full-length cDNA clone is a pivotal step for generating rNDV expressing foreign antigens. Manipulating cDNA clones to express foreign antigens must take several factors into account. The foreign gene of interest should be synthesized as a transcription unit, which comprises a gene start (GS) sequence, the foreign gene, and a gene end sequence (Fig. 2). The transcription unit is then inserted into a noncoding intergenic region of the NDV genome [8]. The number of nucleotides that make up the rNDV genome must follow the “rule of six” to ensure integrity of the genome and efficient virus replication. The insertion site is one of the most important factors affecting efficient expression of the foreign gene. In general, genes closer to the 3' terminus are transcribed more abundantly than downstream genes (the 3' to 5' transcription gradient) due to imperfect restart of transcription at the GS. In other words, the foreign gene is expressed more abundantly and more efficiently when placed closer to the 3' end of the genome [8]. While the foreign gene can be placed at any intergenic site within the NDV genome, the P/M intergenic site is preferred to ensure optimal expression of foreign genes [1617]. The length of the inserted foreign gene also affects efficient expression of the foreign antigen. Incorporating a foreign gene increases the size of a genome and the number of transcriptional units within that genome, which often leads to retarded growth of rNDV in permissive cells [8]. A single NDV vector can accommodate foreign genes up to 4.5 kb in length with a good degree of stability and express at least three different foreign genes [818].
Development of NDV-Vectored Vaccines for Antigen Delivery
NDV vaccine strains infect many mammals but are very safe due to restricted host tropism. Nevertheless, NDV is a strong stimulator of humoral and cellular immune responses at both the local and systemic levels; it also has an adjuvant effect [8]. Importantly, NDV is antigenically distinct from known paramyxoviruses that infect mammals; thus it is unlikely to be affected by pre-existing immunity in mammals [8]. NDV does not establish persistent infection in animals and humans because it replicates only in the cytoplasm [8]. In addition, the rule of six means that genetic changes (e.g., insertions, deletions, and recombinations) affecting the integrity of the genome are rare [8]. Therefore, NDV vaccine strains are potential virus vectors that can be used to develop antigen delivery vaccines for use in animals and/or humans.
Veterinary vaccines
NDV vaccines have been used to develop antigen delivery vaccines for use in cattle/sheep, dogs/cats, pigs, and horses (Table 1). Antigen delivery vaccines for veterinary use include those that protect against bovine herpesvirus 1 (BHV-1) [19], bovine ephemeral fever virus (BEFV) [20], Rift Valley fever virus (RVFV) [21], vesicular stomatitis virus (VSV) [22], canine distemper virus (CDV) [23], rabies virus (RV) [24], Nipah virus (NiV) [25], and West Nile virus (WNV) [26].
BHV-1 is a major cause of respiratory tract disease in cattle. A major problem with the current modified live BHV-1 vaccines is their ability to cause latent infection, with the associated risk of subsequent reactivation [19]. As an alternative strategy, Khattar et al. [19] developed a rNDV vaccine (rLaSota/gDFL) expressing the gD of BHV-1. A single intranasal/intratracheal immunization with rLaSota/gDFL (1.5×107 PFU/dose) induced marked mucosal and systemic antibody responses in calves. Also, intranasal challenge with a high dose of virulent BHV-1 strain Cooper (2×107 PFU) led to a marked reduction in viral shedding and more rapid clearance of challenge virus when compared with mock-infected and challenged controls. These results suggest that NDV may have utility as a vaccine vector that can be used to develop a mucosal vaccine against BHV-1 infection of cattle.
RVFV, a mosquito-borne bunyavirus that infects ruminants (cattle and sheep), causes severe economic losses to the livestock industry and has a high impact on public health in Africa and the Arabian Peninsula [27]. Currently, a live attenuated vaccine (Smithburn strain) and inactivated RVFV vaccines are available; however, although live attenuated vaccines are highly immunogenic, they are not safe for use in livestock [2829]. Inactivated vaccines are safer, but less effective, than live attenuated vaccines and require repeated vaccination [21]. To improve efficacy and safety, Kortekaas et al. [21] developed a NDV-vectored vaccine (NDFL-GnGc) expressing the Gn and Gc glycoproteins of RVFV. Repeat vaccination (two times) of lambs with the NDFL-GnGc vaccine (107.3 TCID50/dose) elicited a high titer of neutralizing antibodies specific for RVFV. These results indicate that NDV has potential as a vaccine vector in sheep.
RV, a rhabdovirus that infects dogs, causes a fatal neurologic disease (rabies) in humans and animals [30]. More than 55,000 people die of rabies each year, with about 95% of deaths occurring in Asia and Africa [31]. Currently available live attenuated RV vaccines induce a weak humoral antibody response, which complicates evaluations of their efficacy [24]. To generate a live attenuated vaccine with improved efficacy, a NDV-vectored rabies vaccine (rL-RVG) containing the RV G glycoprotein was developed [24]. Animal experiments demonstrated that rL-RVG induced production of RV neutralizing antibodies at levels comparable with those induced by a live attenuated RV vaccine (ERA strain) and provided complete protection against challenge with a potentially fatal virus strain (GX/09) for more than 1 year [24].
Human vaccines
NDV vaccine strains have been used as virus vectors to develop antigen delivery vaccines for use in humans (Table 2) [8]. Human NDV vector vaccines under development include those against human immunodeficiency virus 1 (HIV-1) [323334], influenza virus A/H1N1 [25], highly pathogenic avian influenza virus (HPAIV) A/H5N1 [36], severe acute respiratory syndrome coronavirus (SARS-CoV) [37], Ebola virus (EBOV) [38], NiV [25], human norovirus (NoV) [39], respiratory syncytial virus (RSV) [40], and human parainfluenza virus 3 (HPIV-3) [41].
Currently, no live attenuated vaccine is available for HIV-1. However, NDVs have been used as a vaccine vector to deliver the Env and Gag antigens of HIV [323435]. For example, Khattar et al. [34] developed two NDV-vectored HIV vaccines (rNDV-Gag/Env and rNDV-Env/Gag) expressing gp160 Env and p55 Gag, respectively (these genes were inserted between the P and M genes of NDV). Intranasal immunization of guinea pigs with these vaccines induced long-lasting Env- and Gag-specific humoral immune responses. They also induced a marked and efficient cellular and protective immune response in mice after challenge with vaccinia viruses expressing HIV-1 Env and Gag. These results suggest that vaccination with a single NDV vector coexpressing Env and Gag is a promising strategy that increases vaccine immunogenicity and subsequent protective efficacy against HIV.
NDV-vectored vaccines may be useful against human influenza viruses. Nakaya et al. [35] developed a NDV-vectored human influenza vaccine (rNDV/B1-HA) expressing the HA protein of A/WSN/33 influenza virus (H1N1). Two intravenous inoculations of mice with the rNDV/B1-HA vaccine (3×107 PFU/dose) induced a strong anti-influenza virus antibody response and provided complete protection against lethal challenge with influenza A/WSN/33 virus. Recently, DiNapoli et al. [36] developed a NDV-vectored HPAIV vaccine (NDV-HA) expressing the HA protein of A/Vietnam/1203/04(H5N1) HPAIV for use in humans. Its protective efficacy against HPAIV was evaluated in an African green monkey model. Two oro-nasal administrations of NDV-HA (2×107 PFU/dose) induced a high serum titer of HPAIV neutralizing antibodies and a robust mucosal immunoglobulin A response in the respiratory tract. These results suggest that NDV-HA may be an effective human vaccine for HPAIV.
NDV-vectored vaccines can also be used to develop vaccines against deadly emerging viruses. Such viruses include SARS-CoV [31], EBOV [38], and NiV [25]. For example, Kong et al. [25] developed one NDV-vectored NiV vaccine expressing the NiV glycoprotein (rLa-NiVG) and one expressing the F protein (rLa-NiVF), and evaluated their immunogenicity in a pig model. They showed that two intramuscular vaccinations with rLa-NiVF (2×109 EID50/dose) elicited a high titer of NiV-specific neutralizing antibodies (maximum titers of 213) in pigs; these neutralizing antibodies persisted for at least 21 weeks post vaccination, indicating potential utility as a vector vaccine that protects humans and pigs against NiV infection.
NDV can also be used as a vaccine vector to develop vaccines against human viruses for which no whole virus vaccines are available (due to the inability to grow target viruses [e.g., human papillomaviruses, hepatitis C virus, and NoV] in cell culture). For example, Kim et al. [39] developed a NDV-vectored NoV vaccine (a modified rNDV-VP1 vaccine) expressing the capsid protein (VP1) of NoV strain VA387 for human use. The modified rNDV-VP1 grew to a high titer in embryonated chicken eggs (>108 PFU/mL) and produced high levels of VP1 protein in the allantoic fluid. The expressed VP1 protein self-assembled into virus-like particles. Three inoculations of the modified rNDV-VP1 vaccine (3×104 EID50/dose) induced a strong IgG2a-mediated immune response, along with high levels of IFN-γ, tumor necrosis factor-alpha, and interleukin 2 production by splenocytes, in mice; a strong fecal IgA response was also noted. This suggests that the modified rNDV-VP1 vaccine has potential as a live attenuated vaccine against NoV in humans. In addition, NDV-vectored vaccines are also a promising strategy for other human respiratory viruses, including RSV [40] and HPIV-3 [41]. Taken together, these results suggest that a primary vaccination with an NDV-vectored vaccine expressing a foreign protein can be effective. However, the efficacy of an updated or new vaccine based on the NDV vector may be reduced by pre-existing NDV antibodies. This could limit the continuous use of NDV vector vaccines in human.
Development of NDV-Vectored Bivalent Vaccines for Poultry
For several decades, the global poultry industry has used live attenuated or killed vaccines to control and prevent ND. Conventional NDV vaccines are highly immunogenic and induce protective immunity in poultry. Importantly, live attenuated NDV vaccines can be administrated in drinking water, eye drops, and sprays, and even via in ovo injection. This makes it possible to vaccinate poultry (especially young birds) on a large scale. NDV vaccines can be combined with other poultry vaccines (i.e., used as polyvalent vaccines) to control economically important poultry diseases. The production of NDV vaccines is highly cost-effective since they grow to very high titers in embryonated SPF chicken eggs and in cell culture. This makes NDV vaccine strains attractive as viral vectors for the development of polyvalent vaccines against pathogens that could devastate the poultry industry [12].
Highly pathogenic avian influenza vaccines
Global epizootic episodes of H5 HPAIV in wild birds and poultry and their potential to cause a pandemic in humans have driven the development of effective H5 HPAIV vaccines [42], which aim to maintain food security and reduce viral load in the environment. Since the mid-2000s, vaccination programs designed to protect poultry from H5 HPAIV have been implemented by several countries in which the virus is enzootic, including China (and Hong Kong), Egypt, Indonesia, and Vietnam [43]. In general, conventional highly pathogenic avian influenza (HPAI) vaccines for poultry comprise killed virus prepared from a low pathogenic strain or, more recently, from a reverse genetic engineered low pathogenic reassortant virus that derives its hemagglutinin (HA) and neuraminidase genes from a field-virulent virus and its six internal genes from A/Puerto Rico/8/34 (PR8) [44]. Killed avian influenza virus (AIV) vaccines have limitations, including high production costs, poor immunogenicity, and problems with mass vaccination.
Administration of live attenuated AIV vaccines to poultry is strictly prohibited due to the risk of emerging variants arising from reversion to virulence via antigenic drift or antigenic shift in the presence of circulating field viruses [8]. NDV-vectored AIV vaccines are bivalent live attenuated vaccines that are suitable for mass vaccination of poultry on a commercial scale and have a proven safety profile. Basically, NDV-vectored AI vaccines are designed to express the HA protein of AIV, which is critical for protection [4546] (Table 3).
To date, NDV-vectored AIV vaccines have been administered to poultry in China and Mexico. In China, approximately 11.8 billion doses of NDV-vectored AIV vaccine were used to prevent HPAI during 2006–2012 [21]. The HA gene from A/goose/Guangdong/1/1996 (H5N1) was initially incorporated into the NDV vector but was replaced with the HA gene from A/duck/Anhui/1/2006 (H5N1) in 2008, A/duck/Guangdong/S1322/2006 (H5N1) in 2012, and, most recently, with A/chicken/Guizhou/4/2013 (H5N1) [4748]. Approximately 32 million doses of NDV-vectored AI vaccine were used in Mexico (in the form of a live vaccine and a killed vaccine) to prevent H5N2 LPAI in poultry between July 2008 and October 2008. The Mexican vaccine expresses the HA gene from low pathogenic avian influenza virus A/chicken/Mexico/435/2005 (H5N2) [49].
NDV vector vaccines expressing the HA gene of HPAIV show high protective efficacy against HPAIV and virulent NDV in experimental SPF chickens [4550]; however, under field conditions most breeders maintain a high level of immunity to NDV through multiple vaccinations, ensuring that progeny chicks have maternal antibodies to NDV for the first few weeks of life. However, the presence of maternal antibodies could interfere with the efficacy of the NDV-vectored AIV vaccine, resulting in poor antibody responses to NDV and AIV HA. As an alternative strategy, chimeric NDV-vectored vaccines, in which the ectodomains of F and HN proteins are replaced by those of another avian paramyxovirus serotype, have been developed [5156].
Other poultry vaccines
Bivalent NDV-vectored vaccines, which have been developed to prevent diseases of economic importance to the poultry industry, have advantages over traditional vaccines (Table 3). Examples include infectious bursal disease virus (IBDV) [52], infectious bronchitis virus (IBV) [53], infectious laryngotrachitis virus (ILTV) [5455], and avian metapneumovirus (aMPV) [57].
IBDV, a birnavirus that infects chickens, is an important pathogen that causes severe immunosuppression and high mortality in young chickens. Live attenuated vaccines of moderate virulence (especially widely used intermediate plus vaccines) are used widely to prevent infectious bursal disease (IBD); however, they can cause severe side effects (symptoms consistent with IBD) in young chickens. Huang et al. [52] developed a NDV-vectored IBDV vaccine (rLaSota/VP2) expressing the VP2 gene of IBDV, which is responsible for protective immunity against IBDV. The VP2 gene is inserted into the 3'-end non-coding region of the NDV genome. The live IBV vaccine is very safe in young chickens and protects SPF chickens against virulent NDV and virulent IBDV.
IBV, a coronavirus that infects birds, causes respiratory disease and renal disorders (the nephropathogenic strain) in poultry and poor egg production in laying hens worldwide. Currently available live attenuated IBV vaccines risk giving rise to new variants through recombination with field IBVs. This often reduces the efficacy of IBV vaccines. Importantly, live IBV vaccines interfere with the live attenuated NDV vaccine. To overcome the limitations of currently available live vaccines, Toro et al. [53] developed a NDV-vectored IBV vaccine (rLS/IBV.S2) expressing the S2 subunit of the IBV S glycoprotein. Oculo-nasal immunization of chickens (1.0×107 EID50/dose) provided complete protection from clinical disease (mortality) after challenge with a lethal dose of virulent NDV (CA02). The protective efficacy of the rLS/IBV.S2 vaccine was also assessed using a heterotypic protection approach based on priming with a live attenuated IBV Mass-type vaccine followed by boosting with rLS/IBV.S2. The vaccine protected chickens against clinical disease after lethal challenge with a virulent Ark-type IBV strain, leading to a significant reduction in virus shedding when compared with that in unvaccinated/challenged chickens.
ILTV, a herpesvirus that infects birds, causes respiratory disease in chickens. Currently available live attenuated ILTV vaccines are effective, but there are concerns about safety in chickens because of the risks of virulence acquirement and latent infections during bird to bird transmission [5859]. Bivalent NDV-vectored vaccines against ILTV have been developed to overcome side effects associated with the live ILTV vaccine. Kanabagatte Basavarajappa et al. [54] developed a NDV-vectored ILTV vaccine (rNDV gD) expressing glycoprotein D (gD) of ILTV. The protective efficacy of the rNDV gD vaccine against challenge with virulent ILTV and virulent NDV was then evaluated in SPF chickens. Immunizing chickens with rNDV gD (106 TCID50/dose) via the oro-nasal route induced a strong antibody response and provided a high level of protection against subsequent challenge with virulent ILTV and NDV, indicating that rNDV gD has potential as a bivalent vaccine.
Conclusion
NDV vaccine strains show promise as a base from which to develop effective vaccines against pathogens that infect animals and humans. Most NDV-vectored vaccines used for poultry are bivalent and provide protective efficacy against virulent NDVs and several foreign pathogens. NDV-vectored poultry vaccines have been developed to provide protection against HPAIV (A/H5 and A/H7), IBDV, ILTV, IBV, and aMPV. Safe NDV-vectored vaccines have been developed as antigen delivery vaccines for veterinary and human use. Such vaccines express the foreign target antigen and induce robust immune responses at both the local and systemic level as shown with NDV-vectored veterinary vaccines in cattle/sheep (e.g., BHV-1, BEFV, RVFV, and VSV), dogs/cats (e.g., CDV and RV), pigs (e.g., NiV), and horses (e.g., WNV). NDV-vectored human vaccines currently under development aim to provide protection against HIV, HPIV-3, and RSV, newly emerging zoonotic viruses (e.g., HPAIV A/H5, SARS-CoV, EBOV, and NiV), and noncultivable human viruses (e.g., human papillomavirus, hepatitis C virus, and NoV). A primary vaccination by an NDV-vectored vaccine expressing a foreign protein can be effective but the efficacy of an updated or new vaccine based on the NDV vector may be reduced by pre-existing NDV antibodies, which could limit the continuous use of NDV vector vaccines in human.
In conclusion, NDV vaccine strains are attractive vectors that can be used to develop effective vaccines against pathogens that infect animals and/or humans; such vaccines are safe and efficient, and provide high levels of protective immunity, although there is a risk that previous vaccinations can reduce the efficacy of the vectored vaccine.




 XML Download
XML Download