

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Since first human infection with the highly pathogenic influenza virus (HPAI; H5N1) during an outbreak in Hong Kong at 1997, HPAI spread from Asia to Europe and Africa [1]. In 2014, natural infection of HPAI (H5N8) virus containing clade 2.3.4.4 hemagglutinin (HA) have been reported in dogs in South Korea and this virus has been detected in many other countries in Asia and Europe now [2]. During the H5N8 outbreak, two different strains of H5N8 virus which showed highly sequence similarity were isolated from 38 wild birds. H5N8 viruses damaged to more than 200 poultry farms and decreased significant egg production by 60% [3]. Although HPAI (H5N8) has not yet been found to cause disease in humans, there are some subtypes of HPAI such as H5N1, H7N9, and H5N6, which have caused severe illness or death in people. Recently studies showed that vaccines for H5N1 showed cross-reactivity against H5N8 virus [456] but vaccine containing H5N8 viral antigens has not been reported yet.
In general, influenza vaccine was produced in fertilized egg [7]. However, egg-based vaccine production has some limitations such as low productivity of HPAI vaccine and requirement of sealed facilities [8]. To overcome these, a new type of vaccine against influenza, made with virus-like particles (VLPs), has been purposed. Influenza VLPs which are synthesized in cells and released from cells look very like influenza virions. In previous studies, VLPs induced effective immune response more than subunit vaccine composed with recombinant protein antigens [9101112]. Recently, mammalian cell expression systems have been used for production of VLPs and followed by concentration with size exclusion filtration [10]. In this study, we produced influenza H5N8 VLPs for the first time using baculovirus-insect cell expressed system and applied nitrocellulose membrane–based filtration system (NCFS) for concentration of VLP as a vaccine candidate and evaluated yield property and immunogenicity.
Materials and Methods
Plasmid and cloning
H5N8 virus (A/mallard/Korea/W452/2014) derived HA and neuraminidase (NA) gene were generated by polymerase chain reaction (PCR) amplification following each primer: forward BamHI-HA 5′-AGC GGA TCC ATG GAG AAA ATA GTG CTT CTT-3′, reverse XhoI-HA 5′-AGC CTC GAG TTA GAT GCA AAT TCT GCA TTG-3′, forward BamHI-NA 5′-AGC GGA TCC ATG AAT CCA AAT CAG AAA ATA G-3′, reverse XhoI-NA 5′-AGC CTC GAG TTA CAT CTT ATC GAT GTC AAA-3′. HA and NA gene were cloned into pFastBac1 plasmid between BamHI and XhoI. Influenza A/Puerto Rico/8/34 virus (PR8) derived M1 gene was generated by reverse transcriptase polymerase chain reaction and PCR with following primers: forward XbaI-M1 5′-AGG TCT AGA ATG AGT CTT CTA ACC GAG GT-3′ and reverse XhoI-M1 5′-AGG CTC GAG TCA CTT GAA CCG TTG CAT C-3′. M1 gene was cloned into pFastBac1 between XbaI and XhoI. For generation recombinant bacmid, recombinant pFastBac1 were transformed to DH10Bac.
Production of VLPs
Influenza H5N8 VLPs were produced as previously described [13]. Briefly, to generate recombinant baculovirus (rBV) expressing HA, NA, and M1 protein, bacmid encoding HA, NA, and M1 gene were transfected with Spodoptera frugiperda (Sf9) cells by lipofectamin agent (Invitrogen, Carlsbad, CA, USA). HA, NA, and M1 expressed rBV were co-infected with Sf9 cells at a 0.5-1 multiplicity of infection and infected cells were maintained in SF900II medium at 27℃. Day 3 post-infection, culture supernatants were harvested by centrifugation at 4,000 rpm for 15 minutes. Culture supernatants were concentrated by nitrocellulose membrane ultrafiltration system using stirred cell (Amicon, Darmstadt, Germany). Supernatant was filled in cell and gave pressure to cell by N2 gas. Through the membrane with 100 kDa molecular weight cut off, components in culture supernatants less than 100 kDa were filtrated. VLPs in concentrated culture supernatants were purified by ultracentrifugation and performed by 20% and 60% sucrose gradient layer at 30,000 rpm for 1 hour at 4℃. HA titer of each production step were determined by HA assay as previously described [13]. Briefly, serially diluted samples were prepared in 96-well V-bottom plate with phosphate-buffered saline (PBS) buffer and followed by addition of 0.5% chicken red blood cells at 4℃ for 30 minutes.
Characterization of VLPs
Protein concentrations of VLPs were measured by bicinchoninic acid assay kit (Pirece, Rockford, IL, USA). Expression of HA, NA, and M1 in VLPs were determined by western blot analysis, VLPs were separated by 12% polyacrylamide gel and were transferred onto polyvinylidene difluoride membrane. Expression of HA, NA, and M1 were detected by H5N8 virus immunized mice serum or mouse anti-influenza M1 IgG, followed by detection goat anti-mouse IgG horseradish peroxidase (HRP). For negative staining, VLPs were adsorbed on a Formvar-carbon coated EM grid for 1 minute and stained with 2% uranyl acetate for 15 seconds. VLPs were observed by transmission electron microscopy (JEM-1011, JEOL, Tokyo, Japan).
Mice immunization
For animal experiment, six week old of female BALB/c mice (n=6) were immunized intramuscularly with 100 µL of PBS buffer containing 10 µg H5N8 VLPs or immunized intranasally with 30 µL of PBS buffer containing 10 µg H5N8 VLPs. Mice were received twice vaccination at 3 weeks apart. Post 3-week prime or boost vaccination, serum samples were collected by retro-orbital bleeding.
Antibody response
Influenza specific antibody responses in immunized sera were determined by enzyme-linked immunosorbent assay (ELISA) assay as previously described. Briefly, 96-well plates were coated with 100 µL of coating buffer containing H5N8 inactivated virus at 4℃ overnight. Plates were blacked with 200 µL of 3% bovine serum albumin solution at 37℃ for 1.5 hours. Immunized serum were diluted with PBS buffer and added to wells and incubated at 37℃ for 1.5 hours. For determination of H5N8 virus–specific antibody response, plates were incubated with goat anti mouse IgG HRP or goat antimouse IgA at 37℃ for 1 hour and then substrate tetramethylbenzidine solution was used to develop. The optical density was measured at wavelength 450 nm.
Results
Production of influenza VLPs
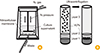
Influenza VLPs expressing HA and NA derived from HPAI (H5N8) were produced by baculovirus-insect cell expression system and concentrated by NCFS ultrafiltration (Fig. 1). In step 1 purification, insect cell culture supernatant co-infected with rBV expressing HA, NA, and M1 influenza proteins were clarified by low speed centrifugation. Total HA activity was about 64,000 hemagglutinating unit (HAU) in 200 mL of clarified insect cell culture supernatant (Table 1). In step 2, clarified cell culture supernatant were put into the stirred cell devices for NCFS and pressurized by N2 gas to force fluid through the nitrocellulose membrane while retaining and concentrating the VLPs (Fig. 2A). The stirred cell used gentle magnetic stirring to control the accumulation of VLPs on the membrane surface under high pressure about 55 psi at room temperature. NCFS was achieved with solution about 10-fold concentration and yielding 80% based on total HA (Table 1). Purity of HA antigen in concentrated solution by NCFS was increased to 6 fold higher than cell culture supernatant based on unit HA titer (Table 1). Finally, VLP solution was purified by ultracentrifugation with 20% and 60% sucrose gradient (Fig. 2B) with 64% recovery yield (Table 1, Fig. 3B). HA titer and unit HA activity in VLP solution were significantly increased during purification process (Fig. 3A, C). As a result, use these NCFS with stirred cell device was effective method to concentrate, diafilter and purify macromolecules such as VLP vaccines.
In vitro characterization of influenza VLPs
Expression of viral antigens in influenza VLPs were confirmed by western blotting analysis. Viral components in influenza VLPs were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis and detected by immune-sera collected from mice immunized with inactivated H5N8 influenza virus (Fig. 4A) and monoclonal anti-M1 antibody (Fig. 4B). HA and M1 proteins were observed in size of 70 and 25 kDa, respectively (Fig. 4A, B). Morphology of influenza VLPs were examined by transmission electron microscopy (Fig. 4C). Influenza VLPs which purified NCFS were identified as nanoparticle with diameter of 80 nm which similar size with influenza virus particles. Therefore, those result indicated that VLPs contained H5N8 influenza viral antigens were effectively produced by baculovirus-insect cell expression system and purified by NCFS with structural similarity.
Humoral immune response induced by influenza VLPs
Influenza VLP vaccines were introduced into group of mice via intramuscular injection or intranasal route twice in three weeks interval (Table 2). Three weeks after boost immunization, mice sera were collected and measured influenza specific antibody responses by ELISA. Both intramuscular and intranasal immunization–induced significant level of virus-specific IgG responses (Fig. 5A). Systemic IgG responses in immunized sera were increase following boost vaccination but secretory IgA responses were not detected in blood even in intranasal immunization (Fig. 5B). Those results indicated that H5N8 influenza VLPs purified by NCFS induced significant level of virus-specific humoral immune responses.
Discussion
In 2014, highly pathogenic influenza A virus (H5N8) was identified in South Korea and widely spread to other country and continent including Asia and Europe. A potential spread of the virus was assumed via the migratory bird routes of duck, geese and swans [3141516]. Although the risk of human infection with HPAI H5N8 is considered very low, vaccine for prevention of HPAI H5N8 should be prepared because it contin-ues to evolve into many genetic lineages and clades. In previously studies, VLP vaccines containing HA and NA derived from H5N1 strain (A/meerkat/Shanghai/SH-1/2012) were showed partial protective immune response against A/MD/Korea/W452/14 H5N8 virus [17] but no any specific VLP vaccines against HPAI H5N8. In this study, we prepared H5N8 VLP vaccine using baculovirus-insect cell expression system. Usually, handling of cell culture medium with large scale is common technical problem during the VLP vaccine manufacturing process. To overcome this, we introduced the NCFS. Estimated size of H5N8 VLP is at least larger than 300 kDa due to their complex compositions such as viral proteins and insect cell membrane. So, VLPs could be concentrated by size exclusion filtration using NCFS stirred cell without significant loss of antigen. After concentration, culture supernatant volume was decrease and HA titer was increased. When we compared total HA unit before and after ultrafiltration with NCFS stirred cell in step 2, HA activity was preserved as approximately 80% and unit HA activity in 1 µg total protein was 6-fold increased. These results were indicated that size exclusion filtration was effective method to concentration VLPs. Finally in step 3, HA titer was increased more than 30 times in comparison with step 1. Total HA unit was preserved as approximately 64% and HA activity in 1 µg total protein was 170 fold increased. These results indicated that VLPs were concentrated and purified successfully by NCFS and sucrose gradient ultrafiltration. Moreover, H5N8 VLPs vaccine induced significant level of IgG antibody both intramuscular and intranasal immunization in mice even after prime vaccination. These results indicated that VLP prepared by NCFS conserved their immunogenicity against antigen. We conclude that these VLP preparation method using nitrocellulose membrane filtration system represent a feasible strategy for VLP vaccine manufacturing.





 XML Download
XML Download