

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Brucellosis is an important zoonotic disease caused by gram-negative facultative intracellular Brucella [1]. According to zoonotic potential and primary host preference, the genus of Brucella is classified into nine species: Brucella melitensis (sheep and goats), Brucella suis (hogs), Brucella abortus (cattle), Brucella ovis (sheep), Brucella canis (dogs), Brucella neotomae (rats), Brucella ceti (cetaceans) and Brucella pinnipedialis (seals), and Brucella microti (common vales) [2]. The zoonotic potential of B. melitensis, B. abortus, and B. suis are considered high and they are most human pathogens [3]. Annually, worldwide incidence of brucellosis is more than 500,000 human infections. Although the disease has a restricted geographic distribution, but it still remains major public health challenge in the Mediterranean region, Asia, Africa, and Latin America. Animal brucellosis causes vast economic losses due to abortion, reproductive failure, and decreased milk production [45]. Among different control measures for the eradication of brucellosis, vaccination is indicated to be the most economic measure for control of the disease in endemic areas [6]. For the development of an effective vaccine, it is necessary to select the best antigen that elicits adequate immune responses (biased towards a Th1) and induces protection against Brucella infection [67].
Bacterial surface exposed antigens are prime vaccine candidates as they represent the initial point of contact between the pathogen and the host immune system. Several Brucella immunogenic antigens have been identified in the outer membrane of this pathogen [689]. According to their molecular mass, the major outer membrane proteins (Omps) of Brucella are classified to group 2 and group 3 proteins representing 36-38 and 25-31 kDa Omps, respectively [89]. The gene products of Brucella major Omps have been designated as Omp2a and Omp2b for the 36-38 kDa porin proteins, Omp25 for the 25-27 kDa Omp, and Omp31 for the 31-34 kDa Omp [9]. Two closely related omp2a and omp2b genes encode and potentially express the 36-38 kDa porin. The genes sequences share about 85% identity and are encoded in the same genetic locus but in opposite directions. The B. abortus biovar1 omp2b gene encodes a 36 kDa protein while omp2a would encode a protein of 33 kDa [9]. The pore-forming activities of Omp2a and Omp2b are different in which B. melitensis 16M Omp2a demonstrates characteristics of a larger pore than Omp2b. Only Omp2b has been detected in Brucella abortus biovar 1, indicating the absence of Omp2a protein at least in this biovar [910].
Recent study focusing on identification of the Brucella protective antigens demonstrated that Omp2b is a priority antigen for designing a recombinant protein vaccine [111]. Moreover, the results of study done by Sung et al. [3] showed that Om2b may be a potential lipopolysaccharide-free protein for developing diagnostic tests. Production of cytokines and nitric oxide (NO) was investigated in RAW 264.7 cells and mouse splenocytes after stimulation with the protein. The production of tumor necrosis factor α, interleukin (IL)-6, and NO were significantly induced in RAW 264.7 cells. Stimulation by Omp2b resulted in significant increase in levels of interferon γ and IL-4 produced by mice splenocytes [3]. Additionally, B. melitensis Omp2b has been identified as anti-apoptotic effector candidate that essentially prevents Bax-induced cell death in Saccharomyces cerevisiae [12].
The aim of the present study was to in silico analysis of Omp2b of the B. abortus 544 as a new vaccine candidate and in vitro expression of the short form of Omp2b (SOmp2b) lacking the signal peptide sequence.
Materials and Methods
Omp2b gene sequence alignment
The full-length sequences of Om2b from several B. melitensis and B. abortus strains were aligned using multiple sequence alignment software (http://workbench.sdsc.edu/) [13]. The identical regions between Omp2b sequences were considered as conserved areas.
Omp31 protein structure analysis
The location of signal peptide and transmembrane regions within the Omp2b sequence were determined using SignalP [14] and TMAP [15] servers, respectively. Surface accessibility, hydrophilicity and antigenicity of the Omp2b protein were determined using Immune Epitope Database (IEDB) analysis resource (http://www.iedb.org). The molecular weight, theoretical isoelectric point (pI), extinction coefficient, total number of positive and negative residues, half-life, instability index, aliphatic index, and grand average hydropathy (GRAVY) of the Omp2b protein were computed using ProtParam server (http://us.expasy.org/tools/protparam.html) [16].
Secondary and tertiary structure prediction
The secondary and tertiary structures of the Omp2b protein were predicted using GOR4 (https://npsa-prabi.ibcp.fr/cgi-bin/npsa) [17] and I-TASSER (http://zhanglab.ccmb.med.umich.edu/I-TASSER) servers, respectively [18].
Validation and analysis of the three dimensional models
The confidence score (C-score), template modeling score (TM-score) and root-mean-square deviation (RMSD) were calculated by I-TASSER server. Energy minimization for the three dimensional (3D) models was performed using Swiss-PDB Viewer 4.1 software. Analysis of the 3D model was made using protein structure analysis (ProSa) server (https://prosa.services.came.sbg.ac.at/prosa.php) [19] and Ramachandran Plot Analysis resource (RAMPAGE) [20]. The Z-score (overall model quality) and energy plots were created by ProSa server.
T-cell epitope prediction
IEDB T-cell epitope prediction tools were used for identification of MHC II epitopes of B. abortus Omp2b protein [21].
Polymerase chain reaction amplification of SOmp2b
Chromosomal DNA of B. abortus strain 544 (Pasteur Institute of Iran) was prepared according to the manufacturer's protocol of DNA extraction kit (Roche, Mannheim, Germany). Polymerase chain reaction (PCR) was performed using forward 5'-CTAGCTAGCATGGCCGACG CAATCGTCGC-3' and reverse 5'-CCGCTCGAGTTAGAACGAACGCTGGAAGCG-3' primers with NheI and XhoI restriction sites (underlined), respectively. The amplification was carried out in a 50 µL total volume component of 500 ng template DNA, 1 µM each primer, 200 µM dNTP-mix, 1× pfu buffer containing MgSO4, and 1 unit of pfu DNA polymerase. PCR conditions used for gene amplification were as follows: hot start at 95℃ for 3 minutes, followed by 30 cycles of denaturation at 94℃ for 45 seconds, annealing at 55℃ for 45 seconds and extension at 72℃ for 1 minute and a final extension at 72℃ for 7 minutes. High pure PCR product purification kit (Roche) was used for purification of the PCR products.
Cloning of SOmp2b gene in pET28a expression vector
The purified SOmp2b PCR product and pET28a expression vector (Novagen, Madison, WI, USA) were digested with NheI and XhoI (Thermo Scientific, Waltahm, MA, USA) restriction enzymes. The ligation reaction has been carried out using T4 DNA ligase (Thermo Scientific) at 4℃, overnight. Transformed E. coli BL21 (DE3) (Stratagene, Kirkland, WA, USA) competent cells were grown on Luria-Bertani agar containing 50 µg/mL of kanamycin (Gibco, Grand Island, NY, USA). Finally, pET28a-SOmp2b positive clones were identified by clone-PCR, restriction enzyme digestion, and sequencing.
Expression and purification of the recombinant SOmp2b
The pET28a-SOmp2b positive clone was cultured in LB broth supplemented with kanamycin (50 µg/mL) in shaker incubator at 37℃ until reached OD at 600 nm of 0.5. Different concentrations of isopropyl β-D-1-thiogalactopyranoside (IPTG; 0.1, 0.2, and 0.4) were added for induction of protein synthesis and the incubation resumed for an additional 4 hours at 37℃. Bacterial cell harvesting was performed by centrifugation at 4,500 rpm for 10 minutes at 4℃. The expressed recombinant protein was purified by Ni-NTA column (Genescript, Piscataway, NJ, USA) under denaturing conditions according to the manufacturer's instructions. Dialyzing of the purified protein was done against 5 M urea, 3 M urea, 1 M urea, and phosphate buffered saline (pH 7.2) at 4℃ overnight.
Sodium dodecyl sulfate polyacrylamide gel electrophoresis and western blotting
Identity of the purified recombinant SOmp2b protein was confirmed by 15% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and western blotting as previously described [24]. The quantity of the recombinant SOmp2b (rSOmp2b) was done by Bradford assay. The purified recombinant protein was stored at -20℃ for further in vivo analysis of its immunogenicity and protective efficacy.
Results
Multiple sequence alignment
Multiple sequence alignment of Omp2b protein sequences demonstrated that the protein sequence is conserved among B. abortus and B. melitensis strains and the alignment score is 99%-100%.
Protein structure analysis
The cleavage site of signal peptidase was between aa22-aa23. Two transmembrane segments aa3-aa28 and aa225-aa253 have been predicted by TMAP tool. The results of surface accessibility, hydrophilicity and antigenicity prediction of the Omp2b protein were summarized in Table 1. The molecular weight and the theoretical pI of the SOmp2b were 36.68 kD and 4.53, respectively. Total numbers of negatively and positively charged residues were 42 and 24, respectively. The half-life of the protein is more than 10 hours in E. coli and in vivo. Aliphatic index, instability index, and GRAVY were 67.12, 24.12 (stable), and -0.294, respectively.
Secondary structure prediction
B. abortus strain 544 Omp2b protein Secondary structure prediction results showed that the protein consist of the percentages of alpha helix, extended strand, and random coil were 19.61, 30.39, and 50, respectively (Fig. 1).
Tertiary structure prediction
In silico prediction of 3D structure of Omp2b was performed using I-TASSER server (Fig. 2A). The C-score (-1.34), TM-score (0.55), and RMSD (9.5) of the model showed that the model has a high confidence value and it is with correct topology. Energy minimization of the best model was done using Swiss PDB Viewer (SPDBV). Structure analysis of the predicted model by Prosa indicated that the model is within the range of native proteins of the similar size (Fig. 2B). Stability of the model was evaluated by the Ranachandran plot (Fig. 2C).
T-Cell epitope prediction
The location of T-cell epitopes with strong affinity for human MHC-II alleles was predicted using IEDB resource. The list of the best human HLA-II epitopes was summarized in Table 2.
Expression and purification of the SOmp2b recombinant protein
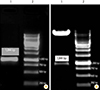
The expression of SOmp2b recombinant protein was induced with 0.1, 0.2, and 0.4 mM IPTG at OD600=0.5 for 4 hours at 37℃. The expected recombinant protein of 36.6 kDa was detected. Purification of the rSOmp2b was done by Ni-NTA affinity chromatography using denaturing method. SDS-PAGE (15%) analyzing of the lysate from the induced E. coli BL21 (Fig. 4A) and the purified protein (Fig. 4B) revealed the expected recombinant protein with the molecular mass of approximately 36.6 kDa. The expression yield of the purified protein was estimated by Bradford assay as 220 µg/mL. The identity of the purified SOmp2b was confirmed by western blotting (Fig. 4C).
Discussion
Selection and production of new vaccine candidates are the primary practical steps toward introducing new vaccines. According to studies published by Jain et al. [1] and He and Xiang [11], Omp2b is an outer membrane protein identified from stationary-phase exoproteome of B. abortus and considered as a potential vaccine candidate against Brucella infection. Bioinformatics analysis indicates that Omp2b is not present in live attenuated B. abortus vaccine strain 19, suggesting that Omp2b probably contributes to the attenuation of this mutant [11]. In the present study, the structure of Omp2b from B. abortus was analyzed using bioinformatics tools. Multiple sequence alignment of different Brucella Omp2b sequences shows the protein is highly conserved among Brucella pathogens (99%-100% sequence similarity) indicating that it could be a new vaccine target against Brucella infection.
Analyzing the physico-chemical parameters of the protein using protparm tools demonstrates that Omp2b is an acidic (pI, 4.53) and stable (with instability index less than 40) protein. The results of Omp2b structure analysis shows that it is a hydrophilic protein regarding its hydropathy values and hydrophilicity prediction of the protein using IEDB tools. Regarding protein secondary structure prediction, Omp2b consists of seven alpha helixes that could stabilize the protein structure. Prediction of the tertiary structure of Omp2b and evaluation of the predicted 3D model using Prosa and Ramachnadran plot indicate that the protein is with the correct topology and within the range of native proteins of the same size.
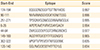
An effective vaccine candidate is one be able to induce strong B-cell and T-cell responses. Therefore, prediction of T-cell and B-cell epitopes are important approaches for designing an optimal vaccine. Prediction of human HLA-II epitopes was performed using IEDB tools and seven best epitopes were listed in Table 2. According to the prediction results, epitope 188NYHIDG YMPDVVGGL202 is the best T-cell epitope with the highest binding affinity score for HLADRB3*01:01. Prediction of continuous B-cell epitopes using BCPred tool resulted in identification of eight epitopes among them 174EQGGDNDGGYTGTTNYHIDG194 epitope was predicted as the best continuous B-cell epitope (score, 0.997). Parker hydrophilicity prediction tool (IEDB resource) also identified this epitope as the most hydrophilic peptide with the maximum score of 7.457. Moreover, all the predicted continuous B-cell epitopes were located in the surface accessible hydrophilic regions of the protein structure. Since strong T-cell and continuous B-cell epitopes confer protection against infection, they could be evaluated experimentally as new anti Brucella vaccine targets. Additionally, identification of conformational B-cell epitopes has been done using Elipro tool. Two potential discontinuous epitopes were predicted that can be used in designing Brucella novel immunodiagnostic tests and antibody production.
In order to in vitro express the SOmp2b, the cleavage site of signal peptidase was predicted SignalP Server and the SOmp2b coding sequence (KP071938) was amplified using specific primers and then cloned in pET28a expression vector. The recombinant protein was successfully expressed upon induction with different concentrations of IPTG and the yield of rSOmp2b was significantly high (220 µg/mL). The expected recombinant protein with the molecular mass of approximately 36.6 kDa was detected by SDA-PAGE and western blotting revealing the identity of the expressed protein.
Our data indicates that Omp2b protein has a potential to induce both B-cell– and T-cell–mediated immune responses and it can be evaluated as a new subunit vaccine candidate against brucellosis. Further studies evaluating the immunogenicity and protective efficacy of the SOmp2b against Brucella pathogens in mice model are underway in our lab.






 XML Download
XML Download