

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Inspite of commendable research efforts from past decades to develop an effective vaccine for tuberculosis (TB), we have still lacked in producing a new vaccine molecule that can surpass safety and imunonogenecity of bacillus Calmette–Guerin (BCG), vaccine for use in humans [1]. As a consequence, which existing TB vaccine continues to cover almost 80% of newborn infants in countries under National childhood immunization programme [2]. Currently World Health Organization (WHO) recommends single dose of BCG vaccine, after birth in countries with high TB incidence rates [3]. Revaccination is not recommended, despite low efficacy of BCG for protection against TB in adults. Vaccination with BCG leads to localized cell-mediated immune response, which subsides within few weeks after vaccination, leaving behind pool of memory cells having immunological memory of primary infection [4].
Circulating antibodies against BCG are thought to be protectitive against TB and provide an initial defense mechanism along with primed T-cell to encounter any primary infection [5]. The developed immune response may vary with age, and depends on various factors like host immunity, level of environmental mycobacterial exposure, and persistent latent TB infection which leads to cross priming and rapid clearance of BCG IgGs [6]. Thus, identifcation of immunological markers in blood of BCG vaccinated and unvaccinated children at different age group may help us to monitor efficacy of BCG protection against TB. This may further help in optimizing the BCG vaccination policies for future, along with implementation of other strategies to boost efficacy of BCG in childrens.
In our earlier studies, we have shown effectiveness of heterologous prime boost regimes to boost level of BCG-induced immunity after primary vaccination [7]. Molecules such as Ag85B peptides showed improved protection in animal studies as against BCG and Ag85B. However, in an effort to develop improved vaccines, and to bridge the gap between animal and human studies, evaluation of such vaccine molecules is needed in peripheral blood mononuclear cells (PBMCs) of individuals with diminished BCG induced immunity after primary vaccination. This ex-vivo approach may be fundamental in terms of evaluating potency of booster dose of Ag85B peptides, BCG, and Ag85B to boost immune response, and offer protection both in animal and humans. Earlier studies by Kashyap et al. [8], in PBMCs of BCG vaccinated children showed that repeated stimulation with BCG boosted initial immunological memory primed by the vaccine. Howver its evaluation in PBMCS of children with low protective levels of BCG along with its comparison with molecules such as subunit and epitope based vaccines was needed to evaluate efficacy of individual candidate molecule to boost waning BCG induced immunity.
The objective of the present was study was (1) study immunological response in serum and PBMCs of children of different age groups vaccinated with BCG vaccine and (2) to study the booster effects of BCG, Ag85B, and Ag85B peptides to boost antimycobacterial immunity in children having diminished BCG-induced response.
Materials and Methods
Vaccines and reagents
BCG vaccine (Moscow strain) was obtained from Serum Institute of India, Pune and stored at 4-8℃ before use. Purified Mycobacterium tuberculosis (Mtb) peptide, Ag85B was obtained from Colorado State University, USA under the TB Research Materials and Vaccine Testing Contract (NO1-AI-75320). Peptides of Ag85B were designed and used as described elsewhere [7]. Freund's incomplete adjuvant was obtained from Genie, Banglore, India.
Study group
Healthy children between 1 month and 18 years vaccinated with and without BCG vaccine were recruited for present study based on predefined inclusion criteria. All participants for the study were recruited from pediatric ward of Govt Medical College and OPD wards of Central India Institute of Medical Sciences. Informed consents were taken from all parents of children after oral explanation about the study. BCG vaccination status in children was determined on basis of BCG scar. In children, whom BCG scar was not evident; BCG vaccination status was asked verbally with his/her kin. Two-three mL blood were collected via venous puncture and distributed equally in sterile plain and ethylenediaminetetraacetic acid (EDTA) vaccutainer tubes. All children recruited for the study were seronegative for human immunodeficiency virus, hepatitis B virus and did not have any other secondary disease.
Follow-up
To evaluate booster effects of BCG, Ag85B, and Ag85 peptides in PBMCs, children with earlier low protective immunological markers, were recruited for present study. Recruitment was made based on earlier records with hospitals. Informed consents were taken from all parents of children after oral explanation about the study. Two or three mL of blood were collected via venous puncture and distributed equally in sterile plain and EDTA vaccutainer tubes.
Serum collection
Blood collected from children was allowed to clot by incubation at 37℃. Serum was separated by centrifugation at 3,000×g for 15 minutes. Collected serum was immediately refrigerated until further use.
PBMC isolation for immunological marker analysis
PBMCs were separated from whole blood of recruited children by density gradient centrifugation using Ficoll Histopaque method as described elsewhere [8]. After counting, the cells were cultured in RPMI-1640 medium keeping the concentration at 2×105 cells/well and were induced with BCG vaccine (100 µL/105 colony-forming unit [CFU]). The cells which were not induced with BCG vaccine were taken as controls. After induction, cells were incubated for 0, 4, 24, 48, 72, 96, and 120 hours in a CO2 incubator. After incubation, cells were centrifuged for 10 minutes at 1,000 rpm. Supernatant obtained was separated and used for anti-BCG IgG titre, adenosine deaminase (ADA), and interferon γ (IFN-γ) estimations.
Evaluation of booster effects of Ag85B peptides, BCG, and Ag85B in PBMCs of BCG vaccinated children
PBMCs were isolated. After counting, the cells were cultured in RPMI-1640 medium keeping the concentration at 2×105 cells/well and were induced with BCG (100 µL). The cells which were not induced were taken as controls. Twelve-hour after incubation, cells were stimulated with BCG vaccine (50 µL/102 CFU), 0.1 mL of Ag85B (25 µg/mL in Freund's adjuvant) and 0.1 mL cocktail of Ag85 peptides (25 µg/mL p1 and p3 in Freund's adjuvant). After incubation for 72 hours, cells were centrifuged for 10 minutes at 1,000 rpm. Supernatant obtained was separated and used for anti-BCG IgG titre, ADA, and IFN-γ estimations by procedure described above.
Estimation of immune parameters
Anti-BCG IgG
Anti-BCG IgG estimation in cell supernatants of PBMCs and serum samples was done using in house developed enzyme linked immunosorbant assay (ELISA) protocol. Briefly the 96-well microtiter plates (MaxisorpImmunoplate, Nalge Nunc International, Naperville, IL, USA) were coated with 10 ng of BCG (diluted in sterile saline). After 3 hours of incubation at 37℃, the plates were washed and blocked with 0.25% bovine serum albumin in phosphate buffered saline (PBS; pH 7.4). After 60 minutes of incubation plates were washed once and kept overnight at 4℃. Next day plates were incubated with cell supernatants/serum samples (1:400 diluted) in PBS. After 45 minutes of incubation plates were washed and incubated with rabbit anti-mouse/goat anti-human IgG, horseradish peroxidase conjugate (1:10,000) for 45 minutes. For color development substrate tetramethyl benzidine in hydrogen peroxide (TMB/H2O2) was added and incubated for 10 minutes. The reaction was stopped by adding 2.5 N sulphuric acid and optical density of color of plates was read at 450 nm.
Analysis of cytokine: IFN-γ
IFN-γ in cell supernatants of PBMCs was measured by an ELISA according to the manufacturer's instructions (Bender Med System, Vienna, Austria) as described elsewhere [8].
Adenosine deaminase
ADA activity in the serum and supernatant was determined at 37℃ according to the method of Guisti and Galanti. Based on the Berthlot reaction, which is the formation of colored indophenol complex from ammonia liberated from adenosine and quantified spectrophotometrically (U.V. visible spectrophotometer, Systronic-Model, Ahmedabad, India). One unit of ADA is defined as the amount of enzyme required to release 1 mmol of ammonia/min from adenosine in standard assay conditions. Results are expressed as units per liter per minute (U/L/min). All assays were performed in triplicates.
Statistical analysis
Data are expressed as mean±standard deviation (SD). Paired t-test was used for obtaining statistical significance. For multiple comparisons, ANOVA was used. Graphs were plotted using sigma plot version 10.0 and Graph Pad Prism 5.0.3 software. p-value of <0.05 was considered statistically significant and p<0.001 for highly significant values.
Results
Study characteristics
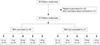
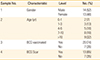
Out of 50 children randomized, 27 were eventually considered for present study (Fig. 1). Baseline characteristics of study population are given in Table 1. Distribution of male and female among recruited children were more or less similar with 52% males (14/27) and 48% females (13/27). Among recruited children, those aged more than 10 years (32%) were the most common as compared to children in age groups 0-1 years (7%), 1-3 years (13%), 4-6 years (18%), and 7-10 years (18%). Out of 27 recruited children, BCG vaccination status was available for 20 children (74%). Among 20 BCG vaccinated children, around 65% (13/20) showed development of BCG scar.
Assessment of immunological markers in blood and PBMC of BCG vaccinated children
Immunological markers were assessed in both blood and PBMCs of recruited children. Based on analysis of immune markers in blood, it was observed that children in age group of 4-6 and 7-10 years showed significantly (p<0.05) increased levels of BCG-specific IgGs which correlated well with T-cell ADA activity and IFN-γ levels. On the contrary, significantly low (vs. other age groups) BCG-induced immunity was observed in children in age group of above 10 years (Fig. 2A). For in vitro studies, isolated PBMCs were stimulated with BCG vaccine and immune response in cell supernatants were analyzed after 48 hours. Based on the assessment of immune response, in vitro studies showed similar results with significantly higher (p<0.001) antimycobacterial immune response (ADA and IFN-γ) in children of 4-6 years, which considerably decreased in children above 10 years of age (Fig. 2B).
Impact of BCG vaccination and scar development on antimycobacterial immunity
To study association of scar development post-vaccination with level of BCG-induced immunity, immunological markers were assessed in blood and PBMCs of BCG vaccinated children with/without BCG scar development. Analysis of biomarkers in blood showed limited impact of BCG scar on observed BCG-induced immunity, with significantly (p<0.05) higher BCG-IgG and T-cell response in children without BCG scar development (Fig. 3A). On the contrary, studies in PBMC model showed, higher BCG-specific IgG and IFN-γ response in cell supernatant of children with BCG scar development (Fig. 3B).
Similarly to study impact of BCG vaccination, all immunological markers were further assessed in BCG vaccinated and non vaccinated children. Similar to above results, it was observed that cell supernatants of BCG vaccinated children were associated with significantly higher anti-BCG IgG and IFN-γ response compared to blood. (Fig. 4A, B). No significant impact of BCG vaccination was observed in blood based on assessment of immunological markers.
Booster effects of BCG, Ag85B, and Ag85B peptides on antimycobacterial immune response in children with low protective cytokine levels
To study the potential of earlier evaluated molecules (BCG, Ag85B, and Ag85B peptides) to boost BCG induced immunity in humans, children with earlier low protective cytokine levels (mostly above 10) were recruited. PBMCs of children were isolated and stimulated with BCG, Ag85B, and AG85B peptides. Fig. 5 shows anti-BCG (Fig. 5A) and cytokine–IFN-γ (Fig. 5B) levels in 72-hour cell supernatants PBMCs of children (n=5) boosted with BCG, Ag85B, and Ag85B peptides. PBMCs of all children showed increased levels of both cytokine and anti-BCG which were comparatively more significant (p<0.001) in cell supernants boosted with BCG and Ag85B peptides compared to native Ag85B (p<0.05).
A comparative analysis of all molecules to boost BCG induced immunity revealed that peptides of Ag85B were able to boost almost 3-fold increase (3±0.2Log2) in IFN-γ expression as compared to earlier levels, followed by booster dose of BCG (2±0.4Log2) and Ag85B (0.8Log2). Cell supernatants of PBMCs boosted with BCG (1.7±0.1) and Ag85B peptides however showed similar fold increase in IgGs to BCG (Fig. 6).
Discussion
Timely assessment and screening of markers of BCG induced immunity is needed to monitor the level of BCG protection post vaccination. Currently, WHO recommends single dose of BCG vaccine immediately after birth for children in high TB endemic regions [9]. Neonatal vaccination is thought to be protective against childhood forms of meningeal and miliary TB [10]; however, many reports are suggestive of waning of the current BCG-induced immunity in children [1112]. This decrease in immunity correlates with increased number of TB cases in later years. Identification of these waning BCG induced protection in children constitutes an important part of health survey programmes and will be helpful to monitor children with high risk for development of TB infection in future. In current study, we intended to study protective efficacy of BCG post-vaccination in children of different age groups with objective to specifically demonstrate course of immunological markers of protection in children vaccinated with BCG with age.
Based on results, it was observed that efficacy of protection post BCG vaccination increases with age, but gradually wanes in later years. Based on the assessment of immunological markers (anti-BCG IgG and IFN-γ) in serum and PBMCs of children vaccinated with BCG, it was found that children in age group of 4-6 years are associated with significantly higher BCG-specific IgG and interferon levels compared to that in age group <10 years. In TB endemic countries like India, cross riming due to environmental mycobacterial infection and persistent latent infection are major reasons for rapid clearance of immunity primed by BCG with age [13]. Our results are therfore in agreement with published reports and suggest that such extrinsic factors may be responsible for reduced efficacy of BCG in later years.
Immune response post-vaccination is dependent on host immunity along with development of BCG scar [14]. Many people post-vaccination are associated with scar development while in some cases, scar may not be observed. To further study impact of scar development on BCG-induced protection; we evaluated protective markers in BCG vaccinated children with/without scar development. Based on results it was observed that BCG scar has limited impact on assessment of markers in blood but has considerable effect in PBMCs of children stimulated with BCG vaccine. The results are suggestive that children with scar have comparatively better developed immunological memory which can be further increased by boosting. Similar results were obtained when immunological markers were analyzed in PBMCs of BCG vaccinated and unvaccinated children, wherein vaccinated children were associated with better immunological recall on stimulation.
The presents results clearly highlights that BCG vaccination, has essential repertoire for short-term protection but lacks development of essential memory cells that may be required to protect against TB disease in later years [15]. Cross priming by environmental mycobacteria along with high persistence of latent TB infection in TB endemic countries like India are main reasons for low efficacy of BCG [1617], therefore booster vaccination with BCG in later years may be required to boost waning BCG-induced immunity in adults for effective protection against TB.
In India, single dose of BCG is given, based on lack of evidences supporting booster concept of BCG. However, revaccination with BCG in adolescence has been routine practice in many countries throughout the world [18]. Revaccination policies adopted in Hungary in late 1950s showed improved protection and sharp decrease in TB cases among purified protein derivative purified protein derivative (PPD) negative individuals and were attributed to the revaccination policies used [19]. Boosting healthy BCG-vaccinated adults in the UK was well tolerated and enhanced PPD-specific CD4+ T-cell responses, but did not induce CD8+ T cells [20]. These studies support our hypothesis that repeated dose of BCG or boosting immunity primed by BCG is needed for effective protection in population where primary effects of vaccination are generally neutralized leaving vaccinated individuals undistinguished from non-vaccinated population.
Another approach to boost BCG induced immunity is use of Mtb antigens and peptides in heterologous prime boost combinations. Heterologous prime boost approach employs sequential administration of vaccines within appropriate intervals by using different antigen-delivery systems such that the immune system is primed to the antigen by using one vector and then boosted with the same antigen delivered through a different vector [21]. Heterologous prime-boost represents a new way of immunization and will stimulate better understanding on the immunological basis of vaccines [22]. The attraction of the heterologous prime-boost approach is that it preferentially expands TB-specific pre-existing memory T cells against antigenic epitopes shared by both the prime and booster vaccines. This approach also minimizes the potential of generating antibody and T-cell neutralizing effect. This approach may also be beneficial for boosting immunity in BCG vaccinated individuals in high TB endemic regions, where primary effect of BCG generally wanes with age due to existing exposure to non tuberculous mycobacteria and persisting latent infection [23].
In our earlier studies in animals, we have shown that heterologous prime boost combination with BCG, Ag85B, and Ag85B peptides increases protective efficacy of BCG vaccine against TB in mice (unpublished data). In an attempt to study protective efficacy of booster doses of BCG, Ag85B, and Ag85B peptides in humans, children with low protective immunological markers post vaccination, were recruited and their PBMCs were boosted all three molecules. PBMCs of children boosted with BCG, Ag85B, and Ag85B peptides showed increased BCG-specific immune reponse compared to stimulated controls. A comparative analysis of booster affects all three candidate molecules revealed that peptides were efficiently able to boost BCG induced immunity in children followed by BCG. No significant impact of Ag85B was observed on per fold rise in BCG-specific response compared to peptides and BCG. Peptide based vaccines offer numerous advantage over subunit vaccines based on Mtb antigens [2425]. The results of present study combined with earlier studies in mice [7] are the thereby suggestive that peptides can be attractive vaccine candidates in near future as they can rapidly stimulate memory cells primed by primary vaccination in neonates and can specifically overcome the limitation of cross priming due to environmental factors in high TB endemic zones. Moreover our studies also highlights a role of booster effect of BCG, which may provide a base for future studies to further explore beneficial effects of booster concept of BCG for protection against TB in childrens and adults in later years. The present approach may be beneficial for evaluation of potential candidate's vaccines prior to trials and may be valuable in predicting the efficacy of vaccines in both preclinical and clinical studies.
The present study was also associated with some limitation. Major limitation was very limited sample size. However considering the study design, it was very difficult to convince participants and their kin to provide blood especially for children less than 5 years. Moreover, due to lack of follow up, we had to exclude lot of subjects which further contributed to low sample size. Authors are aware that such sample size may be difficult to justify the observed conclusion, but results may be taken as preliminary but important observations. We plan to investigate the present study in large cohorts in future.
To conclude, efficacy of protection post-BCG vaccination increases with age, but gradually wanes in later years. Assessment of immunological markers of BCG-induced protection in children suggest that children within age 4-6 years are associated with high immunological markers (anti-BCG IgG and IFN-γ) which eventually diminishes with age thereby suggesting need for booster dose in later years. Assessment of booster effects of BCG, Ag85B, and Ag85B peptides shows, that peptides are attractive candidates to boost immunity in children with low protective immunological markers suggesting they be attractive vaccine candidates to increase protective efficacy of BCG vaccine in high TB endemic regions for protection against TB.





 XML Download
XML Download