

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Rabies causes death in many mammals, including pigs. Over 98% of human rabies comes from dogs [1], and rabies in animals has been transmitted by dogs and various wild carnivores, including foxes, wolves, raccoon dogs, skunks, mongooses, badgers, ferret badgers, and bats, all around the world [2]. Pig rabies is uncommon, representing only 0.1%-1.1% of animal rabies [34]. Nevertheless, there have reports of the identification of rabies in pigs in China, United States, and Brazil [567]. The main clinical signs in a pig bitten by a rabid dog in China are known to be hyperexcitation, roaming, attacks on other pigs, and attempts to jump pen walls to bite pigs [4]. In the United States, clinical signs in pigs with rabies transmitted by wild animals were fever, aggression, restlessness, ptyalism, anorexia, head rubbing, progressive paralysis, depression, and vocalization [6]. Pig rabies results not only in economic losses but also in a serious public health concern. The most effective ways to prevent pig rabies would probably involve blocking the access of wild animals and preparing an effective vaccine.
Animal rabies cases in South Korea have increased in cattle, dogs, and raccoon dogs since 1993. Veterinary authorities made a decision to distribute a rabies bait vaccine to high-risk rabies regions in South Korea as a kind of eradication program [8]. Since 2000, large amounts of bait vaccines have been distributed annually in two provinces, Gyeounggi-do and Gangwon-do, which have been designated as rabies risk regions, with a view to blocking the transmission of animal rabies. The amount of bait vaccine has increased gradually, and it reached 970,000 doses in 2016. In addition to the distribution of a bait vaccine, a new and safe rabies vaccine is needed for domestic pets and wild animals. Previously, we reported on the construction of the recombinant ERAG3G strain containing an Arg-to-Glu amino acid substitution at position 333 of the Evelyn-Rokitnicki-Abelseth (ERA) glycoprotein [9]. The safety and efficacy of the ERAG3G strain was assessed in mice. A single immunization with the ERAG3G strain conferred complete protection in mice and also induced a protective immune response in dogs, cats, and raccoon dogs [910]. Recently, we constructed a new rabies vaccine candidate, the ERAGS strain, which contains two mutations, at positions 194 and 333 of the G protein of ERA strain. The World Health Organization (WHO) recommends that rabies vaccines not cause any adverse symptoms in target and non-target species [1]. The safety of rabies vaccine candidates should be evaluated in rodents, wild animals, and other domestic species [11]. In accordance with this recommendation, the safety and efficacy have already been investigated in mice and raccoon dogs. In this study, we sought to accumulate fundamental data about the safety and immunogenicity of the ERAGS strain in growing pigs and sows, acting as target and non-target animals, and to prepare for the expanded use of the ERAGS strain in pigs susceptible to rabies. Thus, we evaluated whether the new rabies vaccine strain (ERAGS) was safe and whether it induced sufficient immunogenicity in pigs.
Materials and Methods
Cells and viruses
Using a reverse genetics system, a recombinant rabies viruses (RABV), the ERAGS strain, was constructed to serve as a rabies vaccine candidate, as described previously [9]. The ERAGS strain was propagated in murine neuroblastoma (NG108-15) cells maintained in Dulbecco's modified Eagle's medium (DMEM) supplemented with 5% of fetal bovine serum (FBS). The titer of the ERAGS strain was 108.0 FAID50/mL. BHK-21 cells were grown in DMEM with 10% heat-inactivated FBS and antibiotic-antimycotic solution (GenDEPOT, Katy, TX, USA). BHK21 cells and a fixed RABV, CVS11 strain, were used for the fluorescent assay virus-neutralizing (FAVN) test.
Safety and immunogenicity of the ERAGS strain in swine
Four-month-old pigs that were sero-negative against RABV were divided into three groups. Group 1, consisting of six pigs, was inoculated with the ERAGS strain (1 mL, 108.0 FAID50/mL) via the intramuscular (IM) route. Group 2, consisting of six pigs, was administered the same strain and dose via the oral route. Group 3, consisting of three sows, was inoculated with the ERAGS strain at the same dose but using an IM route of administration. Group 4 consisted of three pigs as a control group that received no treatment. All pigs were monitored daily for clinical signs of rabies, such as abnormal behavior, nervous prostration, anxiety, agitation, aggression, and paralysis for 4 weeks' post-administration. Following inoculation, at 2 and 4 weeks, blood was collected from all pigs, including the controls. The serum titers, expressed in International Units per milliliter (IU/mL), were compared with those of a rabies-positive standard serum. The minimum protective titer was determined to be 0.5 IU/mL of FAVN [1]. After finishing the safety and immunogenicity testing in group 3, all sows were euthanized and tissue samples, including the cerebrum, cerebellum, midbrain, spleen, liver, kidney, and lymphoid tissue, were obtained to check for rabies infection.
Serological assay
A virus-neutralizing antibody (VNA) test, the FAVN test, was performed with blood samples from all pigs [1213]. Briefly, a positive reference serum from WHO, adjusted to 0.5 IU/mL, was used as a positive control. Serum samples (50 µL) and the positive and negative controls were distributed in four consecutive wells and then serially diluted three-fold. The RABV (CVS-11 strain) at ~100 FAID50/50 µL was then added to each well. After a 60-minute incubation at 37℃, 50 µL of BHK-21 cell suspension containing 4×105 cells/mL was added to each well, and the microplates were incubated for 72 hours in a humidified incubator with 5% CO2 at 37℃. The cells were fixed in cold acetone (-20℃) for 20 minutes. After three successive washes with phosphate-buffered saline (PBS, pH 7.2), the cells were reacted with a specific monoclonal antibody (Median Diagnostics, Chuncheon, Korea) against the rabies N protein for 45 minutes at 37℃; they were then stained with fluorescein isothiocyanate-conjugated goat-anti mouse IgG+IgM (KPL, Gaithersburg, MD, USA). After washing with PBS, the microplates were air-dried and examined at ×200 using a fluorescence microscope (Nikon, Tokyo, Japan). The titers of serum samples, expressed in IU/mL, were compared with those of the positive standard.
Identification of RABV with a fluorescent antibody test
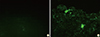
The fluorescent antibody test (FAT) was performed using brain samples (cerebrum, cerebellum, and midbrain) of sows according to the procedure described by the World Organization for Animal Health (OIE) [11]. Smears of brain tissues on slides were fixed in cold acetone (20℃) for 20 minutes. After three successive washes with PBS, the slides were incubated with a monoclonal antibody against RABV for 45 minutes at 37℃ and then stained with the same conjugate mentioned above. After rinsing with PBS, the slides were examined under cover slips at ×200 using a fluorescence microscope. Positive and negative controls were run together with the test samples. Slides showing specific fluorescence were confirmed as positive.
Identification of RABV with reverse transcriptase-polymerase chain reaction
Total RNA was extracted from the homogenates of seven tissue samples (cerebrum, cerebellum, midbrain, spleen, liver, kidney, and lymph node) with an RNA extraction kit (Bioneer, Daejeon, Korea) according to the manufacturer's protocol. The extracted RNA was eluted in 50 µL of RNase- and DNase-free water. Reverse transcriptase-polymerase chain reaction (RT-PCR) was carried out to detect RABV genomic sequences using specific primer sets (RABVDNF and RABVDNR) that amplify the N gene of RABV (Table 1). RT-PCR was performed in a TPersonal 48 thermal cycler (Biometra, Horsham, PA, USA) with a reaction mixture containing 5 µL of denatured RNA, 1 µL of each primer (50 pmol), 5 µL of 5× buffer (12.5 mM MgCl2), 1 µL of dNTP mix, 1 µL of an enzyme mix (reverse transcriptase and Taq polymerase), and 11 µL of distilled water. The cycling program was as follows: cDNA synthesis at 42℃ for 30 minutes, followed by 45 cycles of 95℃ for 15 seconds, 55℃ for 15 seconds, 72℃ for 15 seconds, and a final extension at 72℃ for 5 minutes. The RT-PCR products were visualized by electrophoresis in 1.8% agarose gels containing ethidium bromide. Samples showing a 467 bp band were considered positive.
Statistical analysis
Data are expressed as means±standard deviations. Statistical analyses for VNA titers were conducted with the IBM SPSS version 19.0 for Windows (IBM Corp., Armonk, NY, USA). Differences between groups were analyzed with one-way ANOVA followed by Tukey's post hoc test. For more detailed investigations, the unpaired Student's t-test was individually performed at each time point. A p-value of <0.05 was considered to indicate statistical significance.
Results
Safety and immunogenicity of the ERAGS strain in swine
The growing pigs and sows showed no clinical sign of rabies during the experimental period irrespective of whether the ERAGS was administered orally or via the IM route. As shown in Table 2, unvaccinated pigs also remained in good health during the observation period. As shown in Fig. 1, all growing pigs in group 1 inoculated with the ERAGS strain IM showed VNA titers of 2.6-7.9 IU/mL (mean, 5.92 IU/mL) against RABV at 4 weeks' post-inoculation. However, there were no significant difference in the mean titers in the ERAGS-vaccinated groups of pigs at 2 and 4 weeks after the first vaccination (p<0.05, Tukey's post hoc test) (Fig. 1). Group 2, administered the ERAGS strain orally, showed moderate VNA titers, 0.29-7.9 IU/mL (mean, 2.41 IU/mL). Half (3/6) of group 2 showed protective VNA titers (0.8-7.9 IU/mL) at 2 weeks' post-administration, and four pigs showed protective VNA titers (0.5-7.9 IU/mL) at 4 weeks' post-administration. Two pigs in group 2 showed slight sero-conversion (0.29 IU/mL) but did not reach VNA titers of a protective level. The mean VNA titer of group 2 was higher than those for the sera obtained from the control group (p<0.05, unpaired t-test) (Fig. 1). All sows in group 3, inoculated with ERAGS strain via the IM route, developed protective VNA titers, of 0.8-4.6 IU/mL (mean, 2.7 IU/mL) at 4 weeks' post-inoculation. The three pigs in group 4 remained sero-negative against RABV throughout the test, confirming that no contact transmission occurred between vaccinated animals and the control group.
Detection of RABV in several sow tissues
After performing necropsies on the three sows, tissue samples, including cerebrum, cerebellum, midbrain, spleen, liver, kidney, and lymphoid tissues, were obtained. The brain samples (cerebrum, cerebellum, and midbrain) were subjected to the FAT and RNA was extracted from seven tissue samples. The brain samples obtained from sows inoculated with the ERAGS strain were negative (Fig. 2). The extracted RNAs were subjected to RT-PCR amplifying the N gene of RABV. All samples were negative by RT-PCR (Fig. 3).
Discussion
Before commercialization of a new recombinant rabies vaccine, the safety of the vaccine candidate should be evaluated in target and non-target animals so that all warm-blooded animals, including pigs, that are susceptible to rabies are tested [112]. Domestic pigs may be classified as target animals because an outbreak of pig rabies caused by a rabid dog was reported in China [4]. In our study, high VNA titers were obtained when the ERAGS strain was inoculated into growing pigs and sows via an IM injection. After vaccination with a high dose (108.0 FAID50/mL), six growing pigs and three sows exhibited no clinical sign due to the ERAGS strain. At the time of sacrifice, 35 days following inoculation of the ERAGS strain, clearance of the ERAGS strain from brain samples obtained from the three sows that were inoculated was confirmed by the absence of RABV antigens and gene signals according to FAT or RT-PCR.
The glycoprotein of RABV seems to be important in stimulating a humoral immune response against the rabies vaccine [14]. All growing pigs and sows vaccinated via the IM route developed significant VNA titer levels, ranging from 0.87 to 7.9 IU/mL, indicating that pigs inoculated with the ERAGS strain can be protected against virulent wild RABV, given that WHO considers a 0.5 IU/mL rabies VNA titer to be the minimum protective antibody level in carnivores [1516]. The mean VNA titer of growing pigs was a little higher than that of sows, suggesting that a higher and more rapid VNA antibody response was induced in lighter animals [17].
Since the first outbreak of rabies in Korea was reported in 1907, 16,146 rabies cases have been identified in dogs, cats, cattle, raccoon dogs, horses, and goats [18]. Until 1982, dogs were primarily involved in the transmission of rabies, but, since 1993, raccoon dogs have been responsible for transmission of rabies to other animals in South Korea [19]. Rabies bait vaccine has been distributed as a preventative measure to high-risk rabies regions since 2000.
In the development process of a rabies bait vaccine, the SAD Bern and SAD B19 strains, isolated by successive passaging and simple cloning from the ERA strain, provoked specific mortality in field mice and the meadow vole [20]. A similar study on the safety of the SAG2 strain was conducted in several non-target animals, including wild boar, in areas where bait containing the SAG2 strain was distributed [21]. The results showed that the safety of the SAG2 vaccine was satisfactory. In our study, the oral route was used for growing pigs because this will be the relevant route if wild boar consume bait containing the ERAGS strain. The growing pigs administered a high viral titer of the ERAGS strain via the oral route did not exhibit any clinical signs of rabies during the test period and showed sero-conversion at 4 weeks' post administration. However, 33.3% (2/6) of the pigs did not develop a protective level of antibodies, indicating that pigs need to be administered the virus twice or more. It has been reported that the oral route required more Aujeszky diseases virus vaccine than the intranasal route [22]. Additionally, the presence of antibodies against RABV has been confirmed in wild boar living in Slovenia where anti-rabies vaccination occurred in 2005-2006 [23]. As oral administration of the ERAGS strain in growing pigs did induce an immunogenic response, wild boar consuming the bait may have VNA titers against RABV. Tests of the safety and immunogenicity of the ERAGS strain in target animals are required before commercialization. Thus, further studies concerning the use of the ERAGS as a rabies vaccine will be conducted in pets and domestic animals.
In conclusion, we assessed the safety and immunogenicity of a newly constructed rabies vaccine strain, which was administered via oral and IM routes to growing pigs and sows. No pig inoculated with ERAGS exhibited any clinical sign of rabies over 28 days, and RABV was not detected in tissue samples by FAT or RT-RCR, suggesting that the ERAGS strain may be safe in pigs. Additionally, the vaccinated pigs developed significant VNA titers against RABV, indicating that the ERAGS strain may be immunogenic in swine. Thus, the ERAGS strain is a new, prospective candidate for a rabies vaccine for pigs.



 XML Download
XML Download