

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Salmonella enterica serovar Gallinarum (SG), an intracellular gram-negative bacterium, is a causative agent for a well-recognized septicemic disease of poultry called as fowl typhoid (FT) in many areas, including Latin America, the Middle East, the Pacific Rim, Africa, and Asia. FT is characterized by acute mortality and severe inflammation of internal organs such as liver and spleen and is responsible for significant economic losses to the poultry industry worldwide [12]. A number of attempts have been reported to prevent SG infection by means of vaccination [345678910]. More recently the live SG vaccines have been shown to protect the SG infection in chickens [456]. However, the major safety threats to animal as well as external environment, through the use of live vaccines still remains inexplicable [1112]. Recently, we have successfully demonstrated the use of inactivated SG ghost vaccine to prevent experimentally induced FT infection in chickens at an early age [1314]. The development of bacterial ghosts (BG), produced by protein E mediated lysis under the controlled expression of the phiX174 lysis gene E, is relatively a new approach in order to construct an inactivated vaccine against wide variety of infectious diseases [111516171819]. BG vaccine candidates are considered to maintain all the functional and antigenic determinants in the envelope and are capable of inducing adaptive immune responses [202122232425].
Despite these unusual findings, the optimized immunization strategy to obtain a long lasting immunity to protect chicken from FT is not yet clearly defined. The prime-boost immunization techniques have shown promise for inducing enhanced immune responses against the pathogens [26]. Besides inducing adaptive immunity, the prime-booster vaccinations provide an extra advantage of formation of memory T or B cells, and thus result in the induction of a prolonged humoral and cellular immunity. Upon this vaccination strategy, the humoral immune responses are characterized by enhanced antibody production whereas cell mediated immunity is executed through cytokine mediated activation of macrophages or T-cell subpopulations [27].
A recently constructed safety enhanced inactivated SG ghost vaccine candidate was constructed by using two virulence genes-deleted SG mutant, incorporating an antibiotic resistance free ghost cassette [14]. The present study attempted to optimize the protective efficacy of this candidate and evaluated the immune potential using various prime-booster immunization strategies in chickens.
Materials and Methods
Construction and harvest of SG ghost
Bacterial strains and plasmids used in this study are listed in Table 1. The ghost strain was constructed as described earlier [14]. Briefly, polymerase chain reaction amplification of the ghost cassette was performed using pHCE GAPDH ghost 37SDM as a template and the primers ghost-F-XbaI (5'-TCTAGAGACCAGAACACCTTGCCGATC-3') and ghost-R-XbaI (5'-TCTAGAACATTACATCACTCCTTCCG-3') [12]. The amplified DNA segment was cloned onto the T-vector (Promega, Madison, WI, USA) and was designated pJHL99. The plasmid pJHL101, an antibiotic gene free plasmid containing the ghost cassette, was constructed by enzymatic fusion of pJHL99 and asd + containing pYA3342 plasmids [28]. The safety enhanced SG ghost vaccine was then constructed by transformation of the plasmid pJHL101 into lon, ΔcpxR, and asd deleted JOL967 cells [4] by electroporation and resultant SG transformants were selected on Luria-Bertani (LB) agar plates without diaminopimelic acid (DAP; Sigma, St. Louis, MO, USA). The SG ghost cells were harvested at 18 hours after the lysis induction with temperature upshift from 37℃ to 42℃, washed three times with sterile phosphate buffered saline (PBS), suspended in PBS, and stored at -20℃. All strains were preserved in LB broth with 20% glycerol and stored at -80℃ until use.
Prime-boost immunization in chickens and protection against virulent challenge
The animal experiments described in this study were conducted with approval (CBU 2011-0017) from the Chonbuk National University Animal Ethics Committee in accordance with the guidelines of the Korean Council on Animal Care. One-day-old female Brown Nick chickens were divided into five groups (n=15 per group), and provided with water and antibiotic-free food ad libitum. The chickens were primed with the SG ghosts at 7-days of age and were subsequently boosted at the fifth week of age. Based on the previous preliminary experiment to determine the appropriate dose for inoculation, the oral and intramuscular immunizations with the SG ghost were performed at a concentration of 1×1010 and 1×108 cells/100 µL per chicken, respectively. In the control group A, chickens were orally inoculated with PBS. In group B, the birds were orally primed and boosted with the SG ghost. Group C birds were primed orally and received booster dose by intramuscular route. In group D, the birds were inoculated with prime and booster doses by intramuscular route. Group E birds were received intramuscularly and then received booster dose by oral route. To examine the protection efficacy against the virulent SG challenge, each bird in all the groups was orally inoculated with 100 µL of a suspension containing 1×106 CFU of a wild-type SG JOL394 strain at the eighth week of age. The assessment of mortality and gross lesion scores was carried out as described previously [52930].
Antibody response assessment by indirect enzyme-linked immunosorbent assay
The plasma and intestinal wash samples for weekly determination of IgG and secretory IgA (sIgA) antibody response, respectively, were collected as described previously [3132]. The plasma IgG and intestinal sIgA concentrations were quantified by an indirect enzyme-linked immunosorbent assay (ELISA) against the outer membrane protein (OMP) extracted from the JOL394 SG wild type strain [5] using a chicken IgG and IgA ELISA quantitation kit (Bethyl Laboratories, Montgomery, TX, USA). The wells of a Microlon ELISA plate (Greiner Bio-One GmbH, Frickenhausen, Germany) were coated with 100 µL of OMP at a concentration of 0.2 mg/mL. Wells were incubated with a 1:250 dilution of plasma for 1 hour, followed by incubation with a 1:100,000 dilution of horseradish peroxidase (HRP)-conjugated goat anti-chicken IgG (Bethyl Laboratories) for 1 hour. The bound HRP activity was measured using o-phenylmethylsulfonyl fluoride (Sigma-Aldrich). The optical density at 492 nm was measured with an ELISA reader after the reaction was stopped with 50 µL of 3 M sulfuric acid. The sIgA concentrations were quantified using procedures similar to ones that were used to measure plasma IgG levels.
Antigen specific-lymphocyte stimulation assay
To evaluate the antigen specific lymphocyte proliferation, the lymphocyte proliferation assay in the immunized groups was carried out as previously described [5]. Briefly, peripheral blood mononuclear cells were separated from five chickens randomly selected from each group using a gentle swirl technique [27] at the eighth week of age (day 21 post-booster) and assessed for cell viability using trypan blue dye exclusion test. The viable mononuclear cells added to RPMI-1640 medium supplemented with 10% fetal calf serum, 2 mM L-glutamine, 50 U/mL penicillin, 50 µg/mL streptomycin, and 2 µg/mL fungizone. The 100 µL of suspension (1×105 cells/mL) was incubated in triplicate in the wells of 96-well tissue culture plates with 50 µL of medium alone or medium containing 4 µg/mL of sonicated bacterial cell protein suspension (sbcp) [6] at 40℃ in a humidified 5% CO2 atmosphere for 72 hours. Proliferation of stimulated lymphocytes was measured as previously described [533] .
Assessment of splenic monocyte/macrophages and CD4+ and CD8+ T cells by flow cytometry
To evaluate the immune potential of the ghost vaccine, an independent experiment was carried out to investigate the splenic monocyte/macrophage (M/M) and CD4+ and CD8+ T cells population. The birds were divided into two groups (n=10), group I and group II. In group II, birds received prime and booster dose of the ghost vaccine by intramuscular route at a concentration of 1×108 cells/0.1 mL/chicken at day 7 and the fifth week of age, respectively. The control group I was received intramuscularly with PBS. Five birds from each group were sacrificed at the fourth and eighth week of age. The time point selected for examining the M/M and T-cell population was based on the previous preliminary standardization experiments which was carried out at every week post-prime and -booster immunization (data not shown). The splenocytes were collected and processed as described previously [34]. The M/M population and T-cell markers such as CD3+, CD4+, and CD8+ were examined by flowcytometry. Briefly, the cell suspensions were prepared at a concentration of 1×106 cells/mL in cold PBS. The cells were washed three times in PBS and then incubated with 0.1 mL of appropriately diluted fluorescein isothiocyanate-labeled anti-CD3, biotin-labeled anti-CD4 and phycoerythrin (PE)-labeled anti-CD8a and PE-labeled KUL01 M/M monoclonal antibodies (all SouthernBiotech, Birmingham, AL, USA) in the dark at 4℃ for 30 minutes. The cells incubated with the biotin labeled anti-CD4 antibody were washed three times with cold PBS and were incubated with 0.1 mL of appropriately diluted allophycocyanin-labeled streptavidin (SouthernBiotech) monoclonal antibody in the dark at 4℃ for 30 minutes. After incubation, all samples were washed three times with cold PBS, resuspended in 0.5 mL of PBS, and analyzed with a flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA). Fluorescence-activated cell sorting analysis of 10,000 events was performed using CellQuest software (BD Biosciences).
Quantitative analysis of cytokine mRNA by real-time reverse transcription polymerase chain reaction
Cytokine levels in spleens from the immunized animals were quantified by a quantitative real-time reverse transcription polymerase chain reaction (qRT-PCR) analysis. Splenocytes were harvested as described above and were stimulated with sbcp antigen for 48 hours. Thereafter, total RNA was purified using an RNeasy Mini Kit (Qiagen, Crawley, UK) according to the manufacturer's instructions. Polymerase chain reaction (PCR) primer sets for interferon γ (IFN-γ), interleukin 6 (IL-6) and endogenous positive control GAPDH genes are described in Table 2. The cDNA synthesis was carried out in a 40 µL reaction volume using a TaqMan Reverse Transcription Reagent kit (Applied Biosystems, Warrington, UK), and DNA was amplified and quantified in the ABI Prism 7300 Real-time PCR System (Applied Biosystems) after mixing with SYBR GREEN PCR Master Mix (Applied Biosystems). The qRT-PCR was performed for 40 cycles of 95℃ for 8 seconds, 56℃ for 25 seconds, and 72℃ for 30 seconds.
Statistical analysis
All data are expressed as the mean±standard deviation unless otherwise specified. Analyses were performed with SPSS version 16.0 (SPSS Inc., Chicago, IL, USA). A non-parametric chi-square test was used to analyze significant differences in mortality and gross lesion scores. A one way ANOVA with post hoc Bonferroni adjustments was used to analyze statistical differences in immune responses between the immunized groups and unimmunized control group. Differences were considered to be statistically significant when p-value was ≤0.05.
Results
Prime-boost immunizations and protection against a virulent challenge in chickens
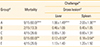
To examine the protective efficacy of the ghost vaccine using prime-booster immunizations, the birds were orally challenged with the wild type SG at the third week post-booster immunization. The birds were subsequently observed for mortality for 14 days post-challenge. All the immunized groups showed significantly lower mortality when compared to the control group A. However, the results displayed in Table 1 indicate that the protection efficacy depended on the different prime-boost immunization strategies. The mortality rate for groups C (primed orally and boosted intramuscularly) and E (intramuscularly primed and boosted orally) was recorded as 26.6% and the birds in group B (orally primed and boosted) showed 13.3% mortality. On the other hand, the birds in group D who received prime-boost immunization by intramuscular route were significantly protected from FT compared to the other group birds (Table 1). Further, the chickens from each group were euthanized for postmortem examination at day 14 post-challenge to validate this protective effect. The birds were evaluated for the presence of lesions in the internal organs such as liver and spleen. All immunized groups demonstrated less organ lesion scores compared to group A. As shown in Table 1, group A demonstrated severe organ lesion scores as 1.66 and 2.00 for liver and spleen, respectively, followed by group E (1.13 and 1.20) and group C (0.93 and 0.80). The group B showed significantly lower lesion scores for liver (0.46) and spleen (0.40) compared to the control group A whereas group D demonstrated significantly lower organ lesion scores compared to all the other groups (Table 1).
Assessment of antibody and lymphocyte stimulation responses in prime-boost immunized groups
Our results indicated that prime-boost immunization with the ghost vaccine using different immunization strategies offered either partial (groups C and E) or significant protection (groups B and D) against the virulent SG challenge. Therefore, we further evaluated the SG antigen specific antibody responses in all the immunized groups by indirect ELISA at each week post-immunization. All the immunized groups showed significantly increased systemic IgG response post-prime immunization and the response was further elevated after booster immunization (Fig. 1). In groups B and C birds, which received prime immunization by oral route, the plasma IgG concentrations were significantly higher than the control group A at second, third, and fourth week post-prime immunization. The IgG response was further increased in groups B and C birds compared to group A after booster immunizations with oral and intramuscular routes, respectively. No significant differences were observed among groups B and C. On the other hand, the birds in groups D and E, which received prime immunization by intramuscular route showed significant IgG response compared to groups A, B, and C at second, third, and fourth week post-prime immunization (PPI). In group E, when birds received a booster dose by oral route, the IgG concentration was continued to remain higher than group A, whereas it did not differ compared to those in groups B and C. In contrast, group D birds demonstrated highly significant IgG response compared to all the other groups after booster. The sequential monitoring of mucosal sIgA antibodies revealed that the intestinal sIgA levels were efficiently induced after oral dose of immunization (Fig. 2). Groups D and E showed significantly higher sIgA response at the first week PPI whereas group B and C showed potent sIgA response at the second and third week PPI compared to the control. After booster immunization, birds in group B showed continuously higher sIgA levels, which differed significantly when compared to the control (Fig. 2). In groups C and D birds, higher sIgA concentration was observed at 5 and 6 weeks PPI whereas group E birds showed higher sIgA response at the seventh week PPI (Fig. 2). The increased sIgA levels in all the groups at the indicated time point significantly differed compared to the control (Fig. 2).
The SG antigen specific lymphocyte proliferative immune responses in all the immunized group chickens were examined by the peripheral mononuclear cell proliferation assay using the specific antigen extracted from the wild type SG. On the third week post-booster immunization, all the immunized birds showed significantly elevated lymphocyte activation responses compared to the control group A (Fig. 3). Group D birds showed significant lymphocyte proliferation response after antigen stimulation compared to all the other groups (Fig. 3). In contrast, no significant differences were observed among groups B, C, and E.
Evaluation of potential of the ghost vaccine to induce cell-mediated immune responses after prime and booster immunization via intramuscular route
As the prime and booster immunization by intramuscular route offered optimized protection against the virulent challenge and induced considerable amount of serum antibody levels in chickens, we further evaluated the immune potential of the ghost vaccine candidate after prime and booster dose of immunization by intramuscular route.
Cellular immune responses in chickens after prime immunization
To examine whether the SG ghost vaccine candidate is capable to induce efficient cell-mediated immune responses, the splenic M/M and CD4+ and CD8+ T-cell subpopulations were investigated in chickens. The flow cytometry analysis revealed that the spleens of the immunized group II showed significant M/M population compared to the un-immunized control group I (Fig. 4A, B). Although group II chickens showed elevated splenic CD4+ and CD8+ T-cell subpopulations there were no significant differences observed between group I and II (p=0.08, data not shown). To correlate the significant M/M population in the spleens of the immunized groups after prime dose of immunization and to investigate the Th1 and Th2 immune responses, we further examined the mRNA levels of IFN-γ and IL-6 cytokines after prime immunization using real-time reverse transcription polymerase chain reaction analysis. The result was expressed as the fold change relative to the average level in the uninfected group. The levels of IFN-γ cytokine were increased in the immunized animals and was >600-fold higher than the unimmunized control at 3 weeks post infection (Fig. 4C), whereas the splenic IL-6 levels of the immunized and un-immunized group did not differ significantly after prime immunization (data not shown).
Booster immunization and cellular immune responses in chickens
Similarly, the splenic populations of M/M and CD4+ and CD8+ T cells were investigated after booster immunization for the immunized and un-immunized chickens. The immunized group II had statistically (p<0.05) higher numbers of splenic CD3+ CD4+, and CD3+ CD8+ T cells compared to the control group (Fig. 5A-C). The M/M population was not significantly different in the group II compared to the control after booster immunization (p=0.1, data not included). The group II also showed 12.5 folds higher levels of IL-6 than that of the control (Fig. 5D) whereas no differences were observed among the groups for IFN-γ mRNA levels (p=0.42, data not shown).
Discussion
The present study attempted to optimize the protective efficacy and immune potential of a newly constructed SG ghost vaccine candidate [14] using various prime-boost immunization strategies. Our results demonstrated that prime-booster immunization with the candidate at day 7 and day 35, respectively, protected chickens against the experimental FT infection (Table 1). The protection efficacy appeared to be dependent upon the prime-boost strategy. Although, there were no significant differences observed among group B (prime-booster with oral route), group C (orally primed and boosted intramuscularly) and group E (primed intramuscularly and boosted orally) for protective efficacy, all three groups differed significantly when compared to group A (non-immunized control). Group D chickens who received prime and booster dose by intramuscular route showed significant protection compared to other groups, evidenced by no mortality and the least organ lesion score. Our data thus may indicate that prime boost immunization strategy via intramuscular route appears to be the optimized strategy to protect chickens against experimental FT infection. Till date, there are no reports available on effective prime boost immunization strategies to prevent FT at an early age. More recently the live SG vaccines have been evaluated in young chickens and showed promising results to protect against virulent SG infection [5635]. Being the live vaccine candidates, all these vaccines may have an advantage of producing long-lasting immunity using a single dose [3637]. However, it is a well known fact that the use of live vaccines has its own boons and banes, latter being the major safety concerns to the animal body and external environment [3839]. Additionally, the commercial live SG9R vaccine is administered at an adult age [40]. Therefore, the present study results show a promising approach to protect chicken against FT infection using prime-boost immunization strategy for young chickens such as 7 days-aged birds, although this new approach using inactivated SG ghost vaccine needs to be validated further.
The correlation between the protective efficacy of SG vaccines and induction of SG antigen-specific adaptive immune responses have been well documented [45681314]. As the prime-boost immunizations with the inactivated SG ghost vaccine offered significant protection, we further investigated the immune responses against the SG antigen in all group's chickens. The sequential monitoring of plasma and intestinal antibodies revealed that the prime-boost immunizations using different routes induced significant rise in plasma IgG concentrations (Fig. 1). Our data suggested that the intramuscular priming induced significantly higher IgG response than the oral priming. The chickens in group C who received prime immunization with oral route showed significant rise in IgG levels when administered with booster dose via intramuscular route. Similarly, group E showed higher IgG concentrations after intramuscular priming and the levels continued to remain higher after oral inoculation with the booster dose. From the data presented in Fig. 1, it appears that the prime-booster inoculation with intramuscular route (group D) induced significantly higher plasma IgG levels compared to all other groups. Indeed, there were no significant differences observed between group D and group E after the first week of booster dose inoculation. However, group D chickens showed significantly higher IgG titer compared to all other groups at the second and third week after booster vaccination. On the other hand, intestinal sIgA levels were significantly higher in orally primed birds compared to birds inoculated with a prime dose via intramuscular route (Fig. 2). The booster dose administration by oral route further elevated the sIgA response in group B chickens whereas group C birds did not show sIgA response upon intramuscular boosting (Fig. 2). The sIgA levels were also significantly increased in group D chickens after booster immunization. Additionally, SG-antigen specific lymphocyte activation responses were investigated after booster immunization and group D chickens showed significantly higher activation responses compared to the other immunized groups, possibly due to the presence of memory cells (Fig. 3). These results thus reinforces the notion that the prime dose of immunization induces adaptive immune responses through activation of B or T lymphocytes, which either leads to antibody production or cellular immunity, and subsequent exposure of booster dose further enhances the immune responses via memory cells which are formed after the prime immunization [26]. Further, our data may also highlight that the prime-booster immunization with either oral route or intramuscular route is capable of inducing desired and more effective immune responses in chickens, as these findings also correlate with the protection data observed for each respective group, group D showing the optimized protection followed by group B and then groups C and E (Table 1).
As our results indicated that the prime-boost immunizations with the ghost vaccine via intramuscular route induced optimized protection, we further evaluated the immune potential of the SG ghost vaccine in chickens. SG are capable of invading the mononuclear macrophage system, therefore cellular immune responses plays a major role in protecting chickens against SG infection [5414243]. The cell-mediated immunity protects the animals against intracellular pathogens by activating cytotoxic and helper T-lymphocytes, macrophages and natural killer cells [21]. These activated T-lymphocytes can act by producing Th1 and Th2 immune responses mediated through various cytokines such as IFN-γ, IL-2, IL-4, and IL-6 or directly destroying the foreign organisms [214445]. In this study, presence of CD4+ and CD8+ T cells and M/M in the spleen, and splenic expression of cytokines such as IFN-γ and IL-6 were investigated in chickens after prime and booster. Our results indicated that the chickens immunized with a prime dose of the SG ghost vaccine via intramuscular route showed significantly higher splenic M/M population and mRNA levels of IFN-γ compared to non-immunized groups (Fig. 4). IFN-γ is the major Th1 cytokine which play an important role in protection against Salmonella infection in avian hosts [44464748]. The most likely mechanism by which IFN-γ exerts its functions in host defense against Salmonella is the activation of macrophages to induce NO production [4950] and MHC class II expression [51]. In the present study, induction of high mRNA levels of IFN-γ along with significantly higher splenic M/M population after prime may suggest that the prime immunization with the inactivated SG ghost vaccine induced mainly Th1 type immune response in the immunized host. Further, the booster immunization induced significantly higher splenic CD4+ and CD8+ T cell population and mRNA levels of IL-6 cytokine in the immunized group chickens (Fig. 5A, B). CD4+ T helper cells (also known as Th cells) play an important role in enhancing the antibody secretion via the production of Th1 cytokines [52] whereas CD8+ cytotoxic T cells play a central role in immune protection in intracellular bacterial parasites such as Salmonella [234446]. The execution of significantly higher splenic CD4+ and CD8+ T cells in the immunized group chickens after booster thus indicate a strong Th1 immune response and correlate with the enhanced antibody production after the booster administration. Additionally, splenic mRNA levels of IL-6 were significantly induced in the immunized chickens after booster (Fig. 5C). IL-6 is a well-known pro-inflammatory cytokine which is produced by Th2 cells and is involved in differentiation of the plasma cells and increased secretory IgA antibody production and thus indicate Th2 immune responses [5354]. The present study results of IL-6 induction in the immunized chickens suggest that the booster immunization induced a potent Th2 type immune response and may also explain that why sIgA levels were significantly elevated after intramuscular booster (Fig. 2, group D). Our results thus indicate that prime immunization with the SG ghost vaccine candidate can induce Th1 type immune response and booster immunization can result in Th1 as well as Th2 immune response in the immunized chickens.
In conclusion, the results of the present study indicate a promising prime-boost immunization approach using the SG ghost vaccine candidate in order to prevent FT in chickens at young age. The results described presently suggests that prime-boost immunization with the candidate via intramuscular route is an optimized immunization strategy and is capable of inducing a potent Th1 and Th2 type immune responses in the immunized host.






 XML Download
XML Download