

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Vaccination is one of the top public health achievements in modern medicine. Majority of the current vaccines are whole cell vaccines [1] which could induce strong immune responses against the complexed form of antigens. Subunit vaccines, in contrast to live-attenuated or inactivated whole cell vaccines, include only specific antigens that stimulate immune responses. Recently, refinement in the fields of science and technology provide chances to develop safer and more maneuverable vaccines [2]. Though entry barriers for licensure are too high, many efforts are focused on the clinical development of the recombinant subunit vaccines [3,4,5]. Despite the advantages of recombinant subunit vaccines, those defined protein products such as recombinant subunit vaccines are less immunogenic compared to older generation whole-cell or conventionally purified vaccines, due in part to the fact that inactivated or attenuated vaccines inherently contain natural pathogen components that serve better immunogens [6,7,8,9]. In this context, effective adjuvants are critically required to help defined vaccines to induce more effective and long-lasting protective immunity.
Adjuvants are vaccine components that potentiate immune responses to an antigen and may modulate them towards more desirable directions [6,7,10,11]. Recent developments in immunology, including the discovery of Toll-like receptors (TLRs) and other innate immune receptors bridging innate and adaptive immunity, have provided new opportunities for the development of more effective adjuvants that could induce more defined protective immune responses. Flagellin, the essential building block of bacterial flagellar filaments, is a pathogen associated molecular pattern that is specifically recognized by the TLR5 on host cell surfaces [12,13]. Previously, we have shown that FlaB, a bacterial flagellar protein from Vibrio vulnificus, binds directly to TLR5 on immune cells and exerts very strong immune reactivity admixed antigens administered through a mucosal route [9,14,15,16]. Since flagellin is a proteinaceous TLR agonist, genetically engineered hybrid vaccines composed of protein antigens and flagellin would be a promising approach for the development of more efficacious adjuvanted subunit vaccines [6,17,18,19]. Tetanus is a lethal medical condition characterized by a spastic contraction of muscles caused by tetanus toxin produced by the Gram-positive bacterium Clostridium tetani. Tetanus can be prevented by neutralizing antibodies against tetanus toxin raised primarily using injectable tetanus toxoid vaccines [20] and efforts to develop mucosally applicable vaccines have not been successful. Several types of combination vaccines (DaP, Tdap, DT, and Td) containing tetanus toxoid as a components are available: D, full-strength doses of diphtheria toxoid; T, tetanus toxoid; P, full-strength doses of pertussis antigens; d, reduced doses of diphtheria toxoid; p, reduced doses of pertussis antigens; a in DTaP and Tdap, acellular. Tetanus toxin fragment C (TTFC), a non-toxic C-terminal fragment of the tetanus toxin, has been used as a recombinant immunogen showing strong protective immunity in animal studies [21,22,23,24,25]. In this study, in a bid to develop a tetanus subunit vaccine applicable through mucosal routes, we hypothesized that FlaB could be a stable fusion partner for TTFC and that the fusion protein would induce efficient protective immunity when applied through mucosal routes. To prove it, we constructed fusion proteins composed of FlaB and TTFC, and tested for anti-tetanus toxin immune potentiating activities. We demonstrate that the mice immunized intranasally with the FlaB-TTFC fusion protein induced significantly higher, compared with TTFC only or the mixture the two proteins, TTFC-specific mucosal IgA and systemic IgG responses and enhanced protective immunity evident by greater survival of immunized mice following challenge with lethal dose of tetanus toxin.
Materials and Methods
Production of FlaB-TTFC or TTFC-FlaB fusion proteins
A DNA fragment encoding the antigenic TTFC polypeptide was polymerase chain reaction-amplified from pTETtac::ttfC [26] using two pairs of primers: F-ttfC-N/R-ttfC-N and F-ttfC-C/R-ttfC-C (Table 1). Similarly, the entire open reading frame of flaB from V. vulnificus CMCP6 was amplified from pCMM250 [9] using two primer pairs: F-ttfC-N/R-ttfC-N and F-ttfC-C/R-ttfC-C (Table 1).
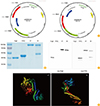
Amplified DNA fragments were initially cloned into the pCR2.1 TOPO vector (Invitrogen, Inc., Carlsbad, CA, USA), yielding plasmids pCMM8209, pCMM8210, pCMM8211, and pCMM8212. DNA fragments were excised using appropriate restriction enzymes and isolated from agarose gels using the QIAEX II gel extraction kit (Qiagen, Hilden, Germany). Plasmid DNA was purified using a QIAprep Spin Miniprep Kit 250 (Qiagen). The 1.3 kb ttfC and 1.1 kb flaB fragments were then cloned into the pTYB12 vector (New England BioLabs, Beverly, MA, USA), yielding plasmids pCMM8213, pCMM8214, pCMM8215, and pCMM8216 (Table 2, Fig. 1A). DNA sequences of the resulting expression vectors were confirmed by the dideoxy-chain termination method. Structure prediction was performed as previously described [28].
Purification of recombinant proteins
Recombinant proteins were prepared as previously described [9,14,15]. Briefly, competent Escherichia coli ER2566 (New England BioLabs) were transformed with pCMM8213, pCMM8214, pCMM8215, and pCMM8216 plasmids by electroporation. Protein expression was induced in mid-log phase cultures by adding 0.4 mM isopropyl-D-thiogalactopyranoside (IPTG). To prepare bacterial lysates for affinity column chromatography, the pellets were resuspended in lysis buffer (20 mM Tris-Cl [pH 7.5], 500 mM NaCl, 1 mM EDTA [pH 8.0], 0.1% Triton X-100, 0.1% Tween 20, 20 M phenylmethylsulfonyl fluoride) and sonicated (Vibra Cell VCX500, Sonics & Materials Inc., Newtown, CT, USA) on an ice. Cell-free sonicate was loaded onto a chitin column and the protein was purified as per manufacturer's instructions. The purity of recombinant proteins was confirmed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), and Western blot analysis with rabbit anti-FlaB and anti-TTFC antibodies. Lipopolysaccharide (LPS) contamination was removed from recombinant proteins using the Affinity Pak Detoxi-Gel Endotoxin Removing gel columns (Pierce Biotechnology Inc., Rockford, IL, USA). Residual LPS content of the protein preparation was determined using the gel-clotting Endosafe LAL kit (Charles River Endosafe, Charleston, SC, USA). The LPS levels in protein preparations were managed to be below the Food and Drug Administration (FDA) guideline (less than 0.15 EU/30 g per mouse). The concentration of obtained proteins was determined by the Bradford dye-binding assay (Bio-Rad Laboratories, Hercules, CA, USA).
TLR5 stimulating activity of the fusion proteins
The TLR5-stimulating activity of the recombinant proteins was determined as previously described [14]. In brief, HEK293T cells were transfected with the p3XFlag-hTLR5 and reporter pNF-κB-Luc plasmids, using Effectene (Qiagen). Twenty-four hours after the transfection, cells were incubated with the purified recombinant proteins for 24 hours. Luciferase activities were normalized to lacZ expression levels using the control expression plasmid pCMV-β-Gal (BD Biosciences Clontech, Palo Alto, CA, USA). Cells were treated with lysis buffer (Promega, Madison, WI, USA), and the luciferase activity was measured by a luminometer (MicroLumat-Plus LB 96V, Berthold, Wilbad, Germany).
Immunization and Ag-specific antibody response
Mice (BALB/c, female, 8-week-old) were immunized under anesthesia (intraperitoneal injection of 100 µL of phosphate buffered saline [PBS] containing 2 mg of ketamine and 0.2 mg of xylazine) three times at 1-week intervals with 20 µL of PBS (negative control group), PBS containing TTFC (1.5 µg), TTFC (1.5 µg) combined with FlaB (1.25 µg) (T+F group), and FlaB-TTFC (2.75 µg) (FT group). One week after the final immunization serum, saliva, vaginal wash, and fecal pellets were collected from various groups to assess Ag-specific antibody responses. All animal experimental procedures were conducted in accordance with the guidelines of the Animal Care and Use Committee of Chonnam National University.
Evaluation of antibody response
To measure the antibody response, enzyme-linked immunosorbent assay (ELISA) was performed as previously described [9]. High binding 96-well ELISA plates (Corning Laboratories, Corning, NY, USA) were coated with 1 µg/mL TTFC protein in PBS (50 µL/well, 4℃, overnight). The plates were washed and blocked with 50 µL/well of a blocking buffer (1× PBS with 0.5% bovine serum albumin, 1 mM EDTA, 0.05% Tween-20) for 1 hour at room temperature (RT). After five washes, serially diluted samples were incubated at RT for 2 hours. Plates were then incubated at RT for 2 hours with horseradish peroxidase-conjugated anti-mouse anti-IgG, -IgG1, -IgG2a, and -IgA Abs (Southern Biotechnology, Birmingham, AL, USA). The color was developed with 50 µL of 3,3',5,5'-tetramethylbenzidine (TMB) substrate (BD Bioscience). The reaction was stopped by adding 50 µL of 1 N H2SO4. The absorbance at 450 nm was measured by a microplate reader (Molecular Devices Corp., Menlo Park, CA, USA). Titers were expressed as the reciprocal log2 values of the highest dilution that yielded an optical density at 450 nm that was ≥0.1 optical density units of negative controls.
Tetanus toxin challenge
To assess the protective immune responses against a lethal challenge, one week after the final immunization, mice were challenged with 600 times LD50 tetanus toxin as described previously [9]. The tetanus toxin (Dongshin Pharmaceuticals, Seoul, Korea) was diluted in 0.2% gelatin PBS and administered subcutaneously to the test mice one week after the last immunization. The mice were observed for 7 days and monitored for paralysis and death as previously described [9,29].
Statistical analysis
All results are expressed as mean±SEM unless otherwise stated. Statistical significance between 2 groups was analyzed using the unpaired Student's t-test. p<0.05 were considered statistically significant. All experiments were repeated three times, and results from representative experiments are shown.
Results
Construction and purification of the fusion proteins
The recombinant proteins were purified by the chitin affinity chromatography. The purity of recombinant proteins was confirmed by SDS-PAGE and the Western blot analysis using specific antibodies against FlaB or TTFC. We confirmed that the fusion proteins were correctly expressed by sequencing the expression vectors. By the SDS-PAGE and Western blot analysis of the fusion proteins, FlaB-TTFC fusion protein was of expected size 92 kDa (Fig. 1B, C). However, the TTFC-FlaB protein expression, by SDS-PAGE and Western blot analyses, presented two major bands of full length and truncated fusion protein (Fig. 1B, C), which suggested that TTFC-FlaB tended to be truncated while the protein was synthesized in E. coli host cells. Interestingly, the truncated protein was detected by either anti-flagellin or anti-TTFC antiserum though its size was similar to FlaB. By the computational comparative three-dimensional modeling [28], the FlaB-TTFC and TTFC-FlaB fusion proteins showed distinct structures (Fig. 1D). The structural analysis demonstrated that FlaB-TTFC had well preserved each (FlaB and TTFC) polypeptide structures and well exposed TLR5 binding domain, while TTFC-FlaB presented a tandem location of FlaB and TTFC structures that may result in defective interaction with TLR5. Hence, the FlaB-TTFC fusion protein was used for further in vitro and in vivo functional tests of the fusion protein.
FlaB-TTFC fusion protein induces strong TLR5 stimulating activity
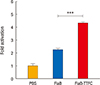
To investigate whether the FlaB-TTFC fusion protein can stimulate TLR5 signaling as efficiently as FlaB, we compared the TLR5-mediated nuclear factor κB (NF-κB) stimulation of the fusion protein by the luciferase reporter assay [14]. The NF-κB-stimulating activity of the FlaB-TTFC fusion protein was compared with that of FlaB only at equimolar concentrations (100 ng of FlaB vs. 219 ng FlaB-TTFC). The result showed that the recombinant protein FlaB-TTFC induced significantly enhanced activation of NF-κB through TLR5 (p=0.0002) compared with FlaB (Fig. 2).
Intranasal administration of FlaB-TTFC induces TTFC-specific antibody responses in systemic and mucosal secretions
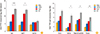
To investigate the levels of TTFC-specific Ab responses induced in systemic and mucosal compartments after immunization with various formulations, serum and mucosal secretions such as saliva, vaginal wash and fecal lysates were prepared from immunized mice one week after the last immunization. Intranasal immunization with FlaB-TTFC fusion protein (FT) resulted in significantly enhanced TTFC-specific serum IgG antibody levels than after TTFC and FlaB mixture (T+F) administration (Fig. 3A). The TTFC-specific IgG1 response in the FT group was significantly higher than that in the T+F group, whereas the IgG2a production profiles were similar between T+F and FT groups (Fig. 3A) suggesting that FlaB had driven the anti-TTFC immune responses preferentially toward Th2 direction. We also assessed TTFC-specific IgA production, the hallmark of mucosal immune responsiveness, in serum and in various mucosal samples. The anti-TTFC IgA titers stimulated by FT were significantly higher in mucosal secretions (p=0.016 in saliva, p=0.011 in vaginal wash, and p=0.003 in feces) than that by the mere mixture of those two proteins (Fig. 3B). These results confirmed that FlaB-adjuvant is an efficient and stable partner for the mucosal TTFC vaccine.
Immune responses induced by intranasal administration of FlaB-TTFC are protective in nature
To test the protective efficacy of the FlaB-TTFC fusion protein, the immunized mice were subcutaneously challenged with 600 times LD50 of tetanus toxin as described previously [9]. After the lethal challenge, survival and paralysis of the mice were monitored for 7 days. All of the vehicle-immunized control mice (PBS) died within 36 hours (Fig. 4A). The final survival percentages of TTFC, T+F groups were 14.3% and 57%, respectively. However, mice immunized with FlaB-TTFC (FT group) showed 100% survival at day 7. Though the statistical significance the survival between T+F and FT was not observed until day 7 (p=0.597; T+F vs. FT by the Kaplan-Meier analysis with log-rank test), signs and symptoms of challenged mice in the two groups were obviously different. The mice in FlaB-TTFC-immunized group looked significantly healthier until day 7 while surviving mice in other groups manifested ill appearance. In this regard, the pathological scores were analyzed as described elsewhere [29]. We gave numerical score as follows: score 0=death within 2 days; score 2=death between 2 and 4 days; score 3=death between 4 and 7 days; score 4=survival on 7th day with marked tetanus; and score 6=survival on seventh day with no tetanus. Groups of mice vaccinated with TTFC alone (TTFC), a mixture of TTFC and FlaB (T+F), and FlaB-TTFC fusion protein (FT) showed 1.14±0.59, 3.86±0.40, and 5.71±0.29 pathological scores, respectively (Fig. 4B). The FT group showed significantly higher pathological score than the T+F group (p=0.0028). The challenge result closely correlated with that of TTFC-specific antibody titer measurement (Fig. 4B).
Discussion
Recombinant vaccine technology is a promising approach to develop safe vaccines inducing specific immune responses [17,18,30]. However, there are several shortcomings that subunit vaccines are inevitably harboring, most important being the low immune-stimulating capacity compared with whole-cell killed vaccines or live attenuated vaccines. Appropriate adjuvant, especially chemically defined ones that could physically associated with the protein subunit vaccines, will compensate the limitations. We have previously suggested that a bacterial flagellin, FlaB from V. vulnificus has strong mucosal adjuvant activity when co-administered with weak mucosal immunogens [9]. Since flagellin is a protein TLR agonist that can easily be engineered by recombinant DNA technologies, a protein-protein fusion approach seems feasible to develop adjuvanted subunit vaccines [14]. We have previously reported that a flagellin serve as a mucosal adjuvant to enhance tetanus toxoid-specific Ab responses in both systemic and mucosal compartments. Furthermore intranasal co-administration of tetanus toxoid and flagellin induce strong protective immunity upon systemic challenge with lethal dose of tetanus toxin [9]. In the present study, we proved FlaB as a stable N-terminal fusion partner of TTFC for mucosal vaccines to induce protective anti-tetanus immune responses. We also observed that the biological activities of fusion proteins significantly vary according to the sequence of fusion. As shown in Fig. 1, FlaB-TTFC fusion protein showed the expected size of 92 kDa while truncated form of TTFC-fusion protein was co-purified with full length TTFC-FlaB. A computational prediction of the three dimensional structure of two proteins indicated that the sequence of fusion could affect the conformation of resulting proteins, which will result in a significant difference in biological activities: i.e., stability, antigenicity, interaction with target immune cells etc. According to the prediction result, FlaB-TTFC protein showed well-preserved domains in both TTFC and FlaB residues and explicit exposure of TLR5 binding domain in FlaB. Depending upon the fusion construction design, two polypeptides will either interfere each other or synergize to induce desirable immune responses. For better biological outcomes, each fused protein should fold stably and should not sterically interfere each other, which emphasized the importance of structural consideration at the very early stage of fusion protein design. In this regard, there should be a robust structural and biochemical studies on the linker residues between fusion proteins in the future.
Estimation of TLR5-dependent NF-κB activity induced by various proteins showed that the FlaB-TTFC fusion protein stimulated the most potent TLR5 signaling. The TF protein induced >2-fold NF-κB activity compared with FlaB only at en equimolar concentration (Fig. 2). These results suggest that C-terminal TTFC fused with N-terminal FlaB retains the TLR5 binding capacity and it interacts with TLR5 signaling apparatus at >2-fold higher efficiency. Since the TLR5 binding domain is hided when flagellin polymers polymerize [12,31], the TTFC polypeptide fused to the C-terminal end of FlaB should have interfered with the assembly of FlaB monomers into polymeric structures and more TLR5 stimulating domains had been exposed. It is also probable that N-terminal FlaB, which is a very well folding stable protein, provides a chance to induce more stable folding downstream TTFC polypeptide and the FlaB-TTFC fusion protein could have longer half-life and consequently have more chance to sustainably stimulate immune cells. This hypothesis could be supported by the in vivo vaccination experiment (Figs. 3, 4). Immunization with fusion protein induced strong antibody responses in all the mucosal compartments owing probably to the commonality of mucosal immune system. Sometimes, intranasally administered vaccines fail to induce efficacious immune responses in the gut. However, the FT fusion protein significantly induced antibody responses even the fecal secretions. Tetanus toxoid, along with mutant tetanus toxin CRM197, is used as a useful conjugation partner of capsular polysaccharide antigens [32]. However, CRM197 is more generally used in commercial vaccines because it does not need further inactivation process after protein purification, which consequently provide ease of standardization and save money required for additional manufacturing process. FlaB-TTFC fusion protein could be considered a useful conjugation partner for low immunogenic epitopes aiming the development of pan-mucosal vaccines.
As a conclusion, the flagellin-fusion protein strategy has been tried by several other groups highlighting the potential utility of flagellin as an adjuvant for inducing a specific immune response [14,33,34,35,36]. Presumably, the FlaB-TTFC fusion proteins will provide a new candidate for the development of polyvalent vaccines such as DT, DTP, and Hib that are applicable through mucosal routes.



 XML Download
XML Download