

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Influenza A virus, a negative-stranded RNA virus that belongs to the Orthomyxoviridae family, is responsible for annual epidemics that cause severe morbidity or death in approximately 5 million people worldwide. The constant pandemic potential of novel influenza subtypes remains a serious threat to public health, as illustrated by the recent pandemics involving swine H1N1 and avian H7N9 [1,2]. Therefore, there is an urgent need to develop effective vaccines against influenza viruses.
The innate immune system, the first line of defense against pathogens, relies on pattern recognition receptors (PRRs) to detect pathogen-associated molecular patterns (PAMPs) [3,4]. For example, influenza genomic RNA is recognized by Toll-like receptor (TLR)-7, which is expressed in late endosomes [5,6], whereas the cytosolic sensor, retinoic acid inducible gene-I (RIG-I), detects 5'-triphosphates within the influenza viral genome in infected cells [7,8]. Signaling via these receptors activates antigen-presenting cells (APCs). These cells produce type I interferons (IFNs) and proinflammatory cytokines, which help to establish an antiviral state, recruit additional immune cells, and direct the adaptive immune response. The type I IFNs (produced mainly by plasmacytoid dendritic cells [DCs]) not only limit viral replication but also act as a natural mucosal adjuvant for intranasally administered influenza vaccines [9]. Many studies show that mucosal immunity induced by natural respiratory influenza infections is more cross-protective against subsequent infection by variant viruses than systemic immunity induced by parenteral immunization with inactivated vaccine [10]. Therefore, to develop an effective vaccine, it is desirable to mimic the process of natural infection that bridges the innate and adaptive immune responses. For example, intranasally inoculated formalin-inactivated influenza virus vaccine induces protective immunity against both homologous and heterologous viruses [11,12]; this is probably because the vaccine retains the viral genomic RNA that stimulates TLR7/8 [13]. By contrast, a split influenza vaccine does not induce antigen-specific immunity when the vaccine is introduced intranasally [14]. However, synthetic double-stranded RNA (dsRNA) can restore the immunogenicity of the vaccine by mimicking PAMPs [14].
Recognition of Viruses by Inflammasomes
Inflammasomes are cytosolic multiprotein complexes that stimulate the activation of caspase-1, which in turn induces the secretion of inflammatory cytokines such as interleukin (IL)-1 beta and IL-18 (Fig. 1) [15,16]. Inflammasome-mediated cytokine release requires two signals: signal 1 (transmitted via TLR, IL-1R, or the tumor necrosis factor receptor) upregulates the expression of pro-1β, pro-IL-18, and nucleotide-binding domain and leucine-rich-repeat-containing protein 3 (NLRP3), whereas signal 2 induces activation of caspase-1 [17]. Thus far, three classes of inflammasome, RIG-I, NLRP3, and pyrin and HIN domain-containing protein (PYHIN), are known to be involved in viral recognition. One study reported that infection with vesicular stomatitis virus or transfection with 5'-triphosphate RNA activates the RIG-I inflammasome [18]. The PYHIN proteins absent in melanoma 2 (AIM2) and gamma IFN-inducible protein 16 (IFI16), the so-called "AIM2-like receptors (ALRs)", bind directly to viral DNA and engage the adaptor protein, apoptosis-associated speck-like protein containing a CARD (ASC) to form the PYHIN inflammasome. The AIM2 inflammasome is activated by intracellular double-stranded DNA (dsDNA) derived from DNA viruses [19-22]. Experiments using AIM2-deficient mice reveal that AIM2 is essential for regulating caspase-1-dependent maturation of IL-1β and IL-18 in response to dsDNA and DNA viruses, such as vaccinia virus and mouse cytomegalovirus [23]. The intranuclear DNA genomes of Kaposi sarcoma-associated herpesvirus, Epstein-Barr virus, or herpes simplex virus 1, are recognized by the DNA sensor, IFI16, which then activates the IFI16 inflammasome [24-26]. In contrast to the RIG-I and PYHIN (ALRs) inflammasomes, which recognize viral nucleic acids, the NLRP3 inflammasome is activated by a wide range of stimuli, including endogenous metabolites, bacterial components, and environmental irritants, in addition to viruses [17]. In the case of influenza virus, the proton-selective ion channel protein, M2, is necessary and sufficient to stimulate the NLRP3 inflammasome pathway, and its localization to the trans-Golgi network is important for NLRP3 activation (Table 1, Fig. 1) [27]. Interestingly, a mutant influenza virus M2 protein, which has lost its proton selectivity and, therefore, enables the transport of other cations (Na+ and K+), mediated increased IL-1β secretion when compared with the wild-type M2 protein [27]. Encephalomyocarditis virus activates the NLRP3 inflammasome via its non-structural protein, 2B, by stimulating Ca2+ flux from intracellular storage sites into the cytosol (Table 1, Fig. 1) [28]. Importantly, 2B proteins expressed by other picornaviruses, poliovirus, and enterovirus 71, also activate the NLRP3 inflammasome. In agreement with Ito et al. [28], Triantafilou et al. [29] showed that the 2B protein of human rhinovirus, another member of the Picornaviridae family, triggers NLRP3 inflammasome activation by inducing Ca2+ flux from the endoplasmic reticulum and Golgi compartments. They also demonstrated that human respiratory syncytial virus (RSV) small hydrophobic protein, which mediates membrane permeability to ions or small molecules [30], is essential for activating the NLRP3 inflammasome (Table 1, Fig. 1) [31]. Recently, Mitoma et al. [32] identified a new pathway that links RNA viruses with NLRP3 inflammasome activation. The DExD/H-box RNA helicase, DHX33, binds to cytosolic dsRNAs in RSV- or reovirus-infected THP-1 cells (a human acute monocytic leukemia cell line) to trigger NLRP3 inflammasome activation (Table 1, Fig. 1). It should be emphasized that the activation status of caspase-1 is different between monocytes (constitutive activation) and macrophages (inducible activation) [33]. Although the cell-type specific differences in caspase-1 activation should be considered in each case, these observations highlight the importance of viroporins, transmembrane pore-forming viral proteins [34], and dsRNA in the virus-induced activation of the NLRP3 inflammasome.
Inflammasomes Control Antiviral Adaptive Immune Responses
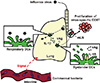
Innate recognition of influenza virus via PRRs not only plays a key role in limiting viral replication at the early stages of infection, but also in initiating antigen-specific adaptive immune responses [3,35]. Recent studies highlight the importance of inflammasome activation for antiviral defense. NLRP3 plays a critical role in limiting lung damage resulting from influenza virus infection [36,37]; however, recognition of influenza virus by the NLRP3 inflammasome does not limit early viral replication in the lung [38]. Using a sub-lethal dose (10 pfu) of influenza virus, the NLRP3-independent inflammasomes activation and its downstream IL-1R signaling events are found to be required to mount adaptive immune responses to influenza virus infection (Fig. 2) [38]. The discrepancies in these results may be explained by the fact that other members of the NLRP family, such as NLRP6 and NLRP12, form an ASC-dependent caspase-1 activating complex [39,40]. So how do inflammasomes activation and downstream cytokine signaling initiate adaptive immune responses? Trans-activation of respiratory DCs via IL-1R is required for the trafficking of antigen-captured DCs to the draining mediastinal lymph nodes (mLNs) and the subsequent induction of virus-specific CD8+ T cell priming (Fig. 2) [41], suggesting that DCs that recognize influenza virus and activate the NLRP inflammasomes are infected by the virus, which renders them unable to perform their antigen-presenting functions in mLNs [42,43]. This should be considered when developing new intranasal vaccines for influenza virus, since heat- or ultraviolet-inactivated influenza virus does not activate inflammasomes [38].
DNA Vaccination and the AIM2 Inflammasome
Because DNA vaccines induce long-lasting humoral and cellular immunity, they are a powerful tool in the fight against infectious diseases. Genetic vaccinations comprise eukaryotic expression plasmids that are inoculated into target cells, which then translate them and express the antigens. The efficacy of this technique correlates with the inflammation induced in muscle cells at the site of DNA vaccination, which causes the release of "danger signals" that induce local inflammatory responses and recruit immune cells to the site of vaccination. In this context, co-administration of plasmid DNA along with adjuvant-like cytokine genes, liposomes, or hyaluronidase, substantially improves the immunogenicity of the DNA vaccine [44-48]. The protective immunity conferred by DNA vaccines has been illustrated using animal models, including those infected by severe acute respiratory syndrome, influenza virus, or human immunodeficiency virus (HIV) [49-51]. DNA vaccines have proved successful in pre-clinical and clinical trials [52-54]. In addition, passive immune-prophylaxis using viral vectors or plasmids encoding neutralizing antibodies induces the long-term expression of antibodies at high concentrations, and provides effective protection against influenza virus or HIV infection [55-58].
AIM2, which recognizes cytosolic dsDNAs of self- and non-self-origin, including viral DNA, combines with the adaptor protein ASC to form a caspase-1-activating inflammasome [59]. A recent report shows that the electrotransfer of plasmid DNA into murine skeletal muscle augments the expression of genes associated with intracellular DNA sensors, including AIM2 [60]. This suggests that the AIM2 inflammasome pathway is required for the immunogenicity of DNA vaccines. In fact, AIM2 knockout mice show a significantly reduced anti-hemagglutinin (HA) antibody response after immunization with a DNA vaccine expressing the HA of influenza virus [61]. Interestingly, blocking IL-1 signaling did not significantly affect the anti-HA antibody responses. Further studies are required to determine the mechanisms by which the AIM2 inflammasome pathway enhances antibody responses after DNA vaccination.
Regulation of Antiviral Immunity by the Intestinal Microbiota
Commensal bacteria are essential for shaping intestinal immune responses [62]; however, the beneficial role of commensal bacteria is not restricted to the intestinal mucosa. Recent studies suggest a link between the intestinal microbiota and antiviral immunity at non-intestinal mucosal surfaces, such as those in the lung. We previously demonstrated that antibiotic-treated mice show impaired adaptive immune responses to influenza virus infection when compared with water-fed mice [63]. Mice treated with antibiotics showed reduced expression of pro-IL-1β, pro-IL-18, and NLRP3 mRNA in the lung, resulting in reduced secretion of mature IL-1β after intranasal infection with influenza virus (Fig. 2). As a result of impaired inflammasome activation in the lungs of antibiotic-treated mice, the number of DCs migrating from the lung to the mLNs was lower than that in water-fed mice after influenza virus infection. Importantly, the administration of a TLR ligand, lipopolysaccharide, was able to restore both the migration of respiratory DCs to mLNs and the virus-specific T cell responses in antibiotic-treated mice. Two follow-up studies identified the molecular mechanism that links the intestinal microbiota to antiviral immunity [64,65]. The induced expression of antiviral defense genes by peritoneal macrophages derived from antibiotic-treated mice, or by splenic DCs derived from germ-free mice, was diminished after stimulation with influenza virus or poly(I:C). This suggests that the microbiota primes APCs by providing tonic type I IFN signals that induce efficient viral recognition and the generation of antiviral adaptive immune responses. Although the main reason of antibiotic treatment to patients infected with influenza virus is to protect the patients from the secondary bacterial infection that causes sever pneumonia, these studies suggest that the administration of antibiotics to patients infected with influenza virus may have negative effects. Although it is still unclear whether it is the species of bacteria or the overall composition of the microbiota that is important for antiviral immunity, the use of gnotobiotic animals and genomic sequencing analysis of the microbiota will highlight new strategies for the development of effective influenza vaccines.


 XML Download
XML Download