

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Rabies is one of the most important zoonoses and caused by rabies virus (RABV), which is mainly transmitted by rabid animal bites, and migrates to the central nerve system and causes fatal encephalitis. Vectors involved in transmitting RABV are dogs, cats, wolves, foxes, skunks, bats, raccoons, and mongooses depending on the countries [1]. Dogs are well known to be the main vector and dog-to-dog or dog-to-other animal transmission is common in many counties including Asia. In addition, raccoons (Procyon lotor) and raccoon dogs (Nyctereutes procynoide) have been involved in RABV circulating in Eastern Europe and the Eastern America since the late 1990s. Raccoon dogs (Nyctereutes procynoide koresis) have been played a key role transmitting rabies to cattle and dogs in Korea [2].
Vaccination is one of the most important tools for prevention and control of rabies in several susceptible animals [3]. National massive vaccination campaigns have lead to dramatic decrease of rabies in countries such as Thailand, India and Korea where numerous of dogs are immunized yearly with rabies vaccines. Nevertheless, rabies still occurs in many countries. Live attenuated vaccine strain, Evelyn-Rokitnicki-Abelseth (ERA) produced in primary porcine kidney cell has been used to immunize dogs, cattle, horses, sheep, and goat and shows protective immunogenicity. But, the ERA strain is not allowed to apply to cats and wild carnivores due to the safety concerns [4]. Vaccination via intramuscular (IM) route is not good enough as there are stray or free-ranging dogs and fierce dogs.
For the prevention of rabies in wild animals, oral immunization with modified live RABV strain, SAD berne, started in 1969 [5]. But, the SAD berne strain had a degree of residual pathogenicity in wild animals and induced a partial immune response in young foxes [6]. The SAD strain was replaced by the Street Alabama-Gif (SAG1) and the first type of rabies bait vaccine strain, SAG2 was developed after successive mutation using anti-glycoprotein monoclonal antibodies. Animal including wild carnivores ingesting ten doses of SAG2 bait remained healthy and showed high rabies neutralizing antibody [7]. The second type of bait vaccine was recombinant adenovirus-vectored vaccines in which both the E1 and E3 gene loci were deleted. The recombinant adenovirus expressing the rabies glycoprotein was developed and distributed in Canada under experimental permit for controlling rabies in dogs, skunks, and raccoons [8]. A canarypox-rabies glycoprotein recombinant vaccine was found to be effective in animals [9].
Other type of bait vaccine is vaccinia-recombinant glycoprotein (V-GR) virus based on Vaccinia virus (Copenhagen strain) recombinated with the rabies glycoprotein gene of the ERA strain. The Copenhagen strain was attenuated from wild Vaccinia virus by replacement of thymidine kinase. A large amount of V-GR vaccine has been distributed for the prevention of rabies in wild foxes and raccoons in European countries and United States since the mid-1990s. The V-GR vaccine has been distributed in rabies risk area of Korea since 2000 [2]. As the V-GR vaccine has helped to prevent any spread of the wild animal RABV in European countries and the United States, the oral rabies vaccination has also contributed to reduction of rabies case in Korea. Nevertheless, the vaccine contains high titer (at least 108.0 TCID50/mL) of a self-replicating orthopoxvirus, which may cause adverse effects in non-target animals including human. Two human vaccinia infections have been reported from V-GR exposure in women with chronic skin disease [10]. Therefore, new alternative bait rabies vaccines applicable to pet dogs and wild animals are needed to eradicate rabies completely. Street strains of RABVs are neuroinvasive and pathogenic in animals, but attenuated strains have a limited ability to invade the central nervous system and can trigger a strong immune response [11]. In this study, recombinant RABVs named as rERA and ERAG3G were constructed using reverse genetic system and the safety and efficacy of the bait vaccine candidate was evaluated in experimental animals.
Materials and Methods
Cells and viruses
The BHK/T7-9 cells, a cloned cell line derived from BHK21 cell [12,13] were maintained in Dulbecco's modified Eagle medium (DMEM) with 10% of tryptose phosphate broth, 5% of heat inactivated fetal bovine serum (FBS) and antibiotics (100 IU/mL penicillin, 10 µg/mL streptomycin, and 0.25 µg/mL (amphotericin B). Hygromycin was added to the medium of BHK/T7-9 cells to make a final concentration of 600 ng/mL. Murine neuroblastoma (NG108-15) cells were maintained in DMEM supplemented with 10% of FBS and were put at 37℃ in 5% CO2 incubator. The attenuated live vaccine strain, ERA was introduced from Canada in 1974 and cloned by several limit dilutions [14]. The ERA strain was used for comparing safety among rERA and ERAG3G strains. The virulent RABV, challenge virus standard (CVSN2c) strain was used for checking efficacy in mice. CVS-11 strain was propagated in BHK21 cells and used for virus neutralizing antibody test.
Construction of recombinant RABV
Full-length cDNA modified into Arg-to-Glu amino acid mutation at position 333 of glycoprotein of the ERA for recombinant ERAG3G strain was cloned into the pUC19 plasmid. Full-length cDNA of ERA strain without modification for rERA was also cloned into the pUC19 plasmid. The nucleocapsid protein (N), phosphoprotein (P) and polymerase, large protein (L) genes obtained from ERA strain were cloned into pT7IRES vector, which possess an internal ribosome entry site. To recover the recombinant RABVs named as rERA and ERAG3G strain from cloned full-length cDNA and helper plasmids, the full-length modified genome plasmid (2 µg) or the full-length genome plasmid (2 µg), EN (0.25 µg), EP (0.02 µg), and EL (0.25 µg) with 9 µL of transfection reagent (Mirus Bio, Madison, WI, USA) were transfected into BHK/T7-9 cells grown in a 24-well tissue culture plate (Fig. 1). After transfection, the cells were screened for cytopathic effects (CPE) for 5 days. The supernatants of the second passage recombinant ERA and recombinant ERAG3G strains were propagated in BHK/T7-9 and NG108-15 cells, respectively and fixed with 80% chilled acetone for 15 minutes. For the staining of the cells, the cells were reacted with specific monoclonal antibody (Median Diagnostics, Chuncheon, Korea) of RABV for 45 minutes, and then stained with fluorescence isothiocyanate (FITC) conjugated goat anti-mouse IgG+IgM. After washing in phosphate buffered saline (PBS), the cells were examined by fluorescent microscopy (Nikon, Tokyo, Japan). After second passage of constructed recombinant RABV the titration of the RABV, was also done by indirect fluorescent assay and the viral titers were determined by 50% fluorescent assay infectious dose per mL (FAID50/mL). In addition, the authenticity of G gene of the rERA and ERAG3G stains was confirmed by direct sequencing of reverse transcription-polymerase chain reaction fragments.
Electron microscopy
The BHK/T7-9 cells inoculated with recombinant RABVs were harvested using a rubber policeman at 72 hours postinoculation. After centrifugation at 1,500 ×g for 10 minutes, the cell pellets were fixed with 2.5% of glutaraldehyde in PBS (pH 7.2) at 4℃ for 2 hours and post-fixed with 1% osmium tetroxide in PBS at 4℃ for 2 hours. After dehydration in a graded series of ethanol and propylene oxide, the cells were embedded in spur resin. Ultrathin sections were made and stained with uranyl acetate and lead citrate, and were observed under an H-7100FA electron microscope (Hitachi Co., Tokyo, Japan).
Safety and potency of RABVs in mice
Four- and six-week-old BALB/c mice were divided into four groups of six animals. Group 1 was inoculated with ERA strain, group 2 with rERA strain, group 3 with ERAG3G, and group 4 remained as control. Each mouse was inoculated with 100 µL or 30 µL of each virus (107.0 FAID50/mL) via IM or intracranial (IC) route. In general, commercial veterinary rabies live vaccines contain about 105.0 FAID50/mL per one dose. Moribund mice were humanely euthanized. Body weight of the mice was measured every three days after inoculation and survival of mice was observed for 15 days after inoculation. To check potency of ERAG3G strain, the 4- and 6-week-old BALB/c mice survived after immunization with ERAG3G strain were challenged by IM injection with a lethal dose of 100 µL of 25 LD50/0.1 mL (CVSN2c strain) into the right leg. Change of body weight in mice and survival of mice were observed for 17 days after challenge.
Safety and immunogenicity of RABV in dogs
The 4-month-old dogs showing seronegative against RABV were divided into two groups. Six dogs were inoculated with ERAG3G strain (1 mL, 108.0 FAID50/mL) via IM route. Two dogs in the control group remained without any treatment. All dogs were monitored daily for adverse effects such as anorexia, prostration, anxiety, agitation, aggression, and paralysis. Following inoculation at 4 weeks, blood was collected from all dogs including control group. The titer of serum samples was expressed in International Units per milliliter (IU/mL) by comparing results obtained with those of the rabies positive standard. The minimum detectable titer was determined as 0.5 IU/mL of neutralizing antibodies according to the World Organisation for Animal Health manual [7].
Serological assay
Virus neutralizing (VN) antibody titer against RABV was determined by the fluorescent antibody virus neutralization test [15]. In brief, a positive reference serum of World Health Organization (WHO) adjusted to 0.5 IU/mL was used as a positive control. Each serum sample as well as the positive and negative controls were distributed in four consecutive wells, and then serially diluted in three-fold. The RABV (CVS-11 strain) containing around 100 FAID50/50 µL was then added to each well. After 60 minutes of incubation at 37℃, a volume of 50 µL of BHK-21 cells suspension containing 4×105 cells/mL was added to each well and the microplates were incubated for 72 hours in a humidified incubator with 5% CO2 at 37℃. The microplates were fixed in cold acetone (-20℃) for 20 minutes. After 3 successive washings with PBS (pH 7.2), the microplates were reacted with specific monoclonal antibody against rabies for 45 minutes at 37℃, and then stained with FITC conjugated goat-anti mouse IgG+IgM. After rinsing with PBS, the microplates were air-dried and were examined at ×100 or ×200 using a fluorescent microscope (Nikon). The titers of serum samples were expressed in IU/mL by comparing results obtained with those of the positive standard.
Results
Construction of recombinant RABVs
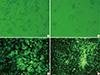
Using the full-length genome mutated or not mutated in glycoprotein gene of ERA strain and helper plasmids, recombinant ERAs named as ERAG3G and rERA strain were constructed in BHK/T7-9 cells successfully. The CPE characterized by rounding and shrinkage was detected after an incubation period of 3 days in BHK/T7-9 cells inoculated with rERA and ERAG3G strains (Fig. 2A, B). The BHK/T7-9 cells inoculated with two strains respectively were fixed with cold acetone and stained with mouse monoclonal antibody against N protein of RABV. The RABV specific fluorescence appeared in the cytoplasm of the infected cell (Fig. 2C, D). The two strains were propagated well in BHK/T7-9 and NG108-15 cells and after two successive passages revealed viral titer of 108.5 FAID50/mL, respectively (data not shown). Electron microscopy revealed that the viral particles were like bullet shape showing typical RABV morphology (Fig. 3A, B). The viral particles of ERAG3G strain were similar to those of rERA strain in size and shape.
ERAG3G strain confers protection from RABV challenge in mice
Body weight in 4- and 6-week-old mice inoculated with ERA, rERA and ERAG3G strains via IM or IC route were measured every three or four days after inoculation. As change of body weight in mice was showed in Fig. 4A and C, average body weight of mice inoculated with ERA and rERA IM decreased sharply at six days after inoculation, and body weight of the mice inoculated with IC decreased faster than IM group (Fig. 4B, D). On the other hand, average body weight in mice inoculated with ERAG3G strain was similar to that of control group. Survival of 4- and 6-week-old mice inoculated with ERA, rERA, and ERAG3G strains via IM or IC route was observed every day after inoculation. All 4-week-old mice inoculated with ERA and rERA strains IM or IC and all 6-week-old mice inoculated with ERA and rERA strains IC respectively died at 12 or 13 days postinoculation. But, only 50% of the 6-week-old mice inoculated with ERA and rERA IM died at 12 days after inoculation. On the contrary, both 4- and 6-week-old mice inoculated with ERAG3G IM or IC and all mice used as control were alive (Fig. 5). The 4- and 6-week-old mice survived after immunization with ERAG3G strain IM or IC were challenged with highly virulent RABV strain CVSN2c on day 15 postimmunization. As shown in Fig. 6, average body weight in mice increased for 17 days after challenge and survival rates did not change for 17 days after challenge. But average weight of control mice challenged with virulent RABV decreased rapidly at seven days post challenge and all mice died at 14 days post challenge. As these results, both ERA and rERA strains were pathogenic to 4- and 6-week-old mice, but ERAG3G strain was non-pathogenic.
Immunogenicity of ERAG3G strain in dogs
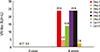
All dogs were sero-negative for rabies VN assay before inoculation and seroconverted 28 days after inoculation. Vaccinated or unvaccinated dogs did not exhibit any clinical signs of rabies during the test. All dogs inoculated with ERAG3G strain IM developed rabies VN titer ranging from 2.62 to 23.9 IU/mL (geometric mean, 15.4 IU/mL) at 28 days after inoculation, but two unvaccinated dogs in group 2 remained sero-negative against RABV throughout the test, confirming that no contact transmission occurred between vaccinees and control (Fig. 7).
Discussion
Rabies has occurred in most Asian countries except Japan, Hong Kong, Malaysia, and Singapore and vector hosts are depending on the countries [16]. In Latin America, vampire bat (Desmodus rotundus) takes responsibility for transmitting the disease to livestock; in Europe, wild animals such as foxes play a key role transmitting the disease; in several countries including India and Thailand, dogs are considered as major vector species [17]. Human rabies is closely associated with animal rabies outbreak and leads to "One Health" concern. The elimination of animal rabies can be achieved by several strategies such as massive vaccination of pets and domestic animals raised within rabies risk regions, implement of program for the management of free ranging dogs and cats and the oral vaccination of wild animals [3]. A large amount of bait vaccines has been distributed for the control and elimination of the rabies in wild animals. Many countries in the world have been carrying out oral vaccination program with SAG2 or V-RG bait. The program has contributed to the elimination of rabies in several countries including Finland, Netherland, and Italy [18].
Several methods for recovering RABV from cloned cDNA have been reported with SAD-B19, RC-HL, and HEP-Flury strains [12,13,19]. It is noted that the efficiency of recovery is various depending on multiple factors including cytomegalovirus promoter activity, cleavage activity of ribozymes, concentration of cDNA plasmids and transfection kit [12]. In fact, recombinant virus is not always produced in the BHK/T7-9 cells, indicating that concentration of full-length cDNA and ratios of three kinds of helper plasmids encoding N, P, and L protein should be optimized for rescuing virus. In this study, two kinds of full-length cDNAs and helper plasmids based on sequence of the ERA strain were prepared and two recombinant ERA strains were rescued in BHK/T7-9 cells. The two ERA strains confirmed by cytophatic effects, fluorescent antibody test and electronic microscopy had similar physical characteristics, suggesting that mutants cannot be distinguished without further analysis such as animal experiment or nucleotide analysis.
It is well known that the amino acid arginine (Arg) at position 333 of the G protein of RABV is responsible for neuro-virulence in animals [4]. If the Arg is replaced by other anino acids such as glutamine, leucine or glycine, viral virulence is greatly decreased [20,21]. And then, it has proven that the ERAG333 strain containing a glutamine at position 333 did not show any pathogenicity in mice older than 7 days [7]. In our study, we identified that the ERAG3G was replaced Arg coding sequence (AGA) into glutamic acid (Glu) coding sequence (GAA) (data not shown). For checking the safety, 4- and 6-week-old mice administered with ERA and rERA strain showing titer of 107.5 and 108.5 FAID50/mL, respectively via IM or IC route revealed pathogenicity at six days after inoculation, whereas mice inoculated with ERAG3G containing 108.5 FAID50/mL were kept in observation for 15 days without displaying any clinical signs. Moreover, mean body weight in the mice did not decrease after inoculation, indicating that the ERAG3G strain is not pathogenic to 4- and 6-week-old mice.
In addition, the efficacy of the ERAG3G strain was evaluated in mice by virulent RABV challenge. Four- or six-week-old mice inoculated with ERAG3G were challenged with 25 LD50/0.03 mL IC and all mice were survived for 17 days. No clinical signs and drop of body weight were recorded in the mice, indicating that single immunization with ERAG3G confers complete protection against the RABV challenge.
As carnivores are primarily responsible for rabies transmission, safety and immunogenicity concerning the oral vaccine candidate should be evaluated in target animals. It has been reported that neutralizing antibodies are crucial protective component against RABV and 0.5 IU/mL of rabies neutralizing antibodies induced by vaccination was considered as the minimum detectable antibody titer in dogs and cats. Cliquet et al. [7] reported that SAG2 strain was safe based on the absence of adverse clinical sign, salivary excretion and absence of replication of the vaccine strain in brain of vaccinated dogs [22]. In this study, we demonstrated that all dogs inoculated with 108.0 FAID50/mL did not show any adverse effects associated with behaviors and all dogs inoculated with ERAG3G developed high neutralizing antibody titers ranging from 2.62 to 23.9 IU/mL at 4 weeks postinoculation, suggesting that dogs inoculated with the ERAG3G strain can be protected by challenge with virulent RABV, which provide the basis to use rabies bait vaccine strain. However, further study concerning usage of oral vaccine strain in accordance with WHO recommendation is needed in dogs and raccoon dogs. In addition, the full-length RABV cDNA containing Pst1 site may be useful as a viral vector to express other foreign gene.
In conclusion, the ERAG3G strain mutated by reverse genetic technology was safe and efficacious in 4- and 6-week-old mice. A single immunization with the ERAG3G strain via IM induced high neutralizing antibody titer against rabies in dogs. Therefore, the ERAG3G strain should be sufficient to allow it use as a live vaccine or oral bait vaccine candidate for domestic and wild animals.





 XML Download
XML Download