

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Influenza A (flu A) viruses belong to the Orthomyxoviridae family of RNA viruses and are the causative agent of influenza, a contagious respiratory viral disease of birds and humans [1]. They possess a negative-sense viral RNA genome packed into 8 individual gene segments which code for at least 11 functional proteins needed for infection, replication, and evasion of antiviral responses. However, recent studies suggest that there could be more viral protein products depending on the virus strain [2,3,4]. Flu A viruses can be further subdivided into distinct subtypes based on the genetic and antigenic properties of their hemagglutinin (HA) and neuraminidase (NA), the envelope glycoproteins express on the surface of virus particles responsible for virus binding to relevant hosts and releasing of newly formed infectious virions, respectively [5]. Sixteen HA and 9 NA subtypes have been detected in wild aquatic birds and poultry throughout the world [1] while new strains (H17-18 and N10-11) have been additionally identified in bat species [6,7].
Generally, flu A viruses are found in migratory waterfowls, the principal natural reservoirs [5]. Part of their genetic variability is due to the absence of a proof-reading mechanism of the flu A viral RNA polymerase leading to faulty replication cycles. Similarly, the segmented nature of the RNA genome is also fundamental to the genetic evolution of flu A viruses. Co-infection of a single host with two or more viruses facilitates genetic reassortment, the rearrangement of genetic material by the exchanging of gene segments, creating progeny viruses with novel gene constellation one of which may have the ability to infect new hosts [1]. Thus, even if they are generally host-specific and not readily transmissible to other species, a wide array of flu A viruses have been found in other animals which include sea mammals, domestic poultry, horses, dogs, pigs, and people [5]. Moreover, flu A viruses are responsible for the annual global influenza epidemics and occasional pandemics in human populations. In recent years, number of flu A viruses crossing the animal-human host species barrier have been increasing. This article provides a general overview of these zoonotic infections and the constant threat of a looming pandemic.
Variant Avian Flu A Viruses Occasionally Breach the Animal-Human Interface
Avian influenza viruses (AIVs) recognize specific sialic acid receptors of susceptible cells lining the intestinal gut of avian hosts [8]. However, specific modifications in HA through adaptation shifts receptor recognition, which in turn effectively switches binding preferences to specific receptors expressed by mammalian hosts, including humans. Further genetic alterations at the HA cleavage motif of some subtypes (e.g., H5 and H7) could make HA readily cleaved by ubiquitous cellular proteases [9]. Accumulation of polybasic residues on this site, typically seen among highly pathogenic avian influenza (HPAI) H5 and H7 viruses, promotes efficient biological activation facilitating growth in various organs outside the respiratory tract [9,10]. Other viral factors are also found in the remaining gene segments which altogether promote propagation at different temperatures, efficient replication of the viral genome, attenuation of host immune responses, and resistance to antiviral drugs [11]. Collectively, these features account for the virulence, expansion of host range, and interspecies transmission of AIVs.
Various AIV subtypes have occasionally crossed the animal-human interface commonly occurring after close contact to infected birds (Table 1). Of note, humans also possess AIV-susceptible cells in the lower respiratory tract (e.g., lungs) apart from mammalian-specific receptors; such receptors are not expressed by upper respiratory tract tissues (e.g., nasal passages and trachea) [12]. Thus, it is not surprising that extensive or prolonged exposure to affected poultry increases the chances of interspecies transmission.
Among the vast pool of AIVs in nature, HPAI A/H5N1 viruses have been considered to present the greatest concern to cause the next flu pandemic (Table 1). Since its first appearance in Guangdong, China in 1996, it has already spawned various clusters of antigenically divergent HPAI H5 lineages that spread widely in Asia towards Europe and Africa [13], causing devastating poultry outbreaks along its path. In humans, more than 640 HPAI A/H5N1 infections worldwide have been recorded since 1997 by the World Health Organization wherein about 60% were fatal [14]. Aberrant innate immune responses have been shown to be a hallmark of the atypical virulence of the HPAI A/H5N1 virus that results in the high incidence of mortality [15,16,17]. In 2014 alone, new cases have been already reported in Asia with Vietnam and Cambodia recording fatalities for the year [14]. The first imported case in North America was also recently confirmed in a resident of Alberta, Canada who returned from Beijing, China in December 27, 2013, was hospitalized, and died after few days due to viral infection [18]. At present, there are no imminent indications that contemporary HPAI A/H5N1 viruses have the capacity to support efficient viral transmission in humans. However, two independent studies have simultaneously demonstrated how few changes in the HPAI A/H5N1 viral genome could confer airborne transmission between mammals [19,20].
Human infection with A/H9N2 AIVs was first documented in Hong Kong in 1999 [21]. Retrospectively, it was postulated that the HPAI A/H5N1 virus, which caused the 1997 human outbreaks also in Hong Kong, acquired most of its gene segments from avian A/H9N2 subtypes that were circulating in 1997 [22]. Sporadic cases of AIV A/H9N2 zoonoses followed ever since with at least 16 human total infections as shown in the influenza virus sequence database (http://www.ncbi.nlm.nih.gov/genomes/FLU/FLU.html) [23], including a recent Hong Kong case in late December 2013, the first in four years [24]. Serologic studies also showed that some poultry workers in China and India were positive for A/H9N2 virus infection through HA inhibition and virus micro-neutralization assays [25,26,27]. Notably, avian A/H9N2 infections only induced relatively milder clinical disease with no mortality in contrast to its more virulent HPAI A/H5N1 cousin.
Prior to 1996, human infections with the H7 AIV subtype have been primarily due to laboratory or occupational exposures [28]. Direct avian-to-human transmission of a low pathogenic avian influenza (LPAI) A/H7N7 was first confirmed in 1996 on an English woman while tending to her apparently infected pet ducks [29]. In 2002, LPAI A/H7N2 virus was detected in Virginia [30] and isolated in an immunocompromised New York resident [31]; this subtype was also found among poultry workers in Wales, England [32]. Similarly, an LPAI A/H7N3 was recovered from a poultry attendant in the UK [33] whereas retrospective serologic analysis of workers who responded during the 2002-2003 H7N3 Italian poultry outbreak identified 7 persons seropositive [34]. In contrast to these LPAI H7 strains, HPAI A/H7N3 variants were responsible for zoonotic infections among poultry workers in British Columbia, Canada [35] and Jialisco, Mexico [36]. Before 2013, the largest human H7 outbreak occurred in the Netherlands [37] caused by an HPAI A/H7N7 tallying over 80 cases including one fatality. Up to that point, all of the H7 patients have only presented generally mild respiratory disease commonly accompanied by conjunctivitis, except for the United States cases [28]. However, pathobiology of H7 AIVs changed when human cases caused by an A/H7N9 virus emerged since late March 2013 killing a quarter of those infected [38]. Unlike its A/H5N1 and A/H9N2 relatives, A/H7N9 does not induce remarkable influenza-like disease signs in afflicted poultry albeit efficient replication [39,40]. The pathogen, a product of multiple reassortment between strains found in migratory birds and prevailing A/H9N2 viruses in Chinese poultry [41], does not contain the pathotypic HA cleavage motif but show indications that it can infect and replicate well in mammalian species [39,40]. As of writing, more than 350 cases have been confirmed with over 70 casualties mostly from mainland China. Imported cases in Hong Kong, Taiwan and a Chinese visitor in Malaysia are the only other reports outside the Chinese territory [42], indicating that it hasn't yet gone global dissemination. Notably though, human infections with A/H7N9 viruses have been characterized by rapid progression to severe pulmonary disease and acute respiratory distress syndrome, reminiscent of human HPAI A/H5N1 disease [39,40,41]. The scale of these zoonotic infections also highlights the need for enhanced surveillance in neighboring countries. The geographic location of South Korea puts it well within the East Asian flyway of migratory waterfowls. In addition, exchange of tourists between China and Korea are constantly growing due to close proximity and improved air travel. Thus, A/H7N9 could be carried over by infected migratory wild birds and/or human traveler/visitors as what happened in the imported case of Malaysia.
Apart from the more popular strains above, H6 and H10 AIV infections have been also recorded. In May 2013, the first human case of A/H6N1 AIV infection was reported [43]. The woman patient, who in worked in a deli shop and has not visited or had any contact with poultry, was hospitalized after developing high fever, cough and shortness of breath, but fully recovered upon early medication. Occurrences of A/H10N7 infections were also separately recorded in Egypt [44] and Australia [45]. Although those patients had minor symptoms and eventually recovered, two of three A/H10N8 AIV infections in China since December 2013 succumbed to death due to infection [46,47]. Like its A/H7N9 predecessor, A/H10N8 does not cause remarkable disease in birds and harbor segments from A/H9N2 virus segments in Chinese poultry [46].
Pigs as Intermediate Hosts that Could Hasten Virus Evolution
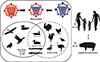
With the inherent capacity of flu A viruses to evolve and adapt, it is only a matter of time before novel AIV variants capable of sustaining person-to-person transmission arise. The role of pigs as a focal point in the generation of novel flu A viruses with pandemic potential has been constantly highlighted. The presence of mixed populations of sialic acid viral receptors that line the pig respiratory tract epithelia make pigs susceptible to avian and human flu strains, apart from their own viruses [8,48]. Hence, infection of pigs with AIVs could facilitate adaptation to mammalian hosts particularly humans via accumulation of adaptive mutations (Fig. 1). Meanwhile, simultaneous infection of a pig with at least two viruses from different sources and lineage could promote genetic reassortment.
Through natural and experimental conditions, some of the notable AIVs mentioned above have the capacity to infect and become established in swine populations. Although domestic pigs appear to have low susceptibility [49], HPAI A/H5N1 viruses have been isolated from pig herds in China [50,51,52,53] and Indonesia [54,55]. Serologic exposure to HPAI A/H5N1 viruses was also detected in asymptomatic pigs in Vietnam [56]. Since 1998, several A/H9N2 viruses have been also isolated from pigs in Hong Kong and other provinces of China [56,57,58,59]. Somehow, a more recent study predicted that A/H9N2 viruses may have become endemic among domestic pigs [59]. Variants of the A/H7N9 virus have been also shown to productively infect pigs in experimental settings but were unable to transmit to other animals [60]. It is also noteworthy that the internal gene constellations of the newly emergent A/H7N9 and A/H10N8 viruses contain corresponding segments from A/H9N2 virus subtypes prevailing in Chinese poultry [38,40,46]. Given the general reputation of pigs with regards to virus ecology and evolution, their role in the genesis in either or both of these AIVs that are making ripples for pandemic concerns cannot be ruled out. Regardless, introduction of these AIVs in swine populations could provide alternative means to hasten genetic evolution (Fig. 1).
Pre-pandemic Vaccine Development and the Need for Novel Antiviral Drugs
Vaccination remains the most effective countermeasure against the spread of novel and potentially pandemic flu A viruses [61]. For the past decade, several vaccine strategies and preparations for humans have been developed and are being tested ranging from inactivated, live-attenuated, recombinant protein-based, virus-like particles to DNA-based and adenovirus-vectored vaccines [62]. Since the world's population is expected to have no pre-existing immunity against A/H5N1 strains, the use of efficacious and safe adjuvants to maximize vaccine coverage is also being considered and evaluated. There are also increasing efforts in developing universal vaccines that target conserved domains of various viral proteins [62]. Acknowledging the probability of a future A/H5N1 pandemic, the World Health Organization Global Influenza Programme for its part has recommended the development of representative pre-pandemic H5N1 vaccine candidate viruses that represent antigenically and genetically distinct H5 strains, particularly those that have infected humans through contact with ill or dead infected birds, for stockpiling [63]. Such pre-pandemic vaccines have been prepared by reverse genetics, and safety tested prior to distribution for pilot vaccine production that may be used for experimental studies and for stockpiling [64].
Because A/H7N9 infection of humans is very rare, it is not surprising to find that no pre-existing immunity was detected against the A/H7N9 virus in any of the age groups tested in serologic assays [65]. Apparently, receipt of current seasonal human flu vaccines did not also induce detectable cross-reactive antibodies suggesting that even current seasonal vaccination will not afford protection. In a more promising note though, few groups have shown that some H7-specific vaccine preparations, developed by applying strategies learned from the H5 and 2009 pandemic vaccine development efforts, could offer protection from A/H7N9 virus challenge in animal models [66,67,68,69]. However, their efficacy in human subjects remains to be evaluated. Although pre-pandemic H5 and H7 vaccines are in the pipeline for development, there is also a dire need for other subtype-specific candidate vaccines (e.g., H6, H9, and H10). Furthermore, strategies that can augment vaccine coverage will be invaluable in the event of a pandemic to meet global demands.
Antiviral drugs provide an important alternative public health intervention for preventing and treating influenza when a vaccine is not available. Currently, two major classes of anti-flu drugs are available which inhibit the release of infectious viruses (NA inhibitors) and block the matrix protein 2 ion-channel (adamantane derivatives), respectively [70,71]. However, given the plasticity of the flu A viral genome, the emergence of drug-resistant strains against these classes of prophylactic agents could arise. Apart from that, the timing of treatment is also significant on their efficacy. Notably, emerging variants of the human-infecting A/H7N9 virus already gained NA inhibitor resistance without losing their viability and pathogenicity [72]. Altogether, these underscore the need for the development of new antiviral drugs and other novel preventive and therapeutic measures.
Conclusion
At present, none of the zoonotic AIVs above indicated efficient human-to-human dissemination beyond index cases. However, the growing number of cases sparks continuing worries that they could eventually evolve into variants that will cause global concern. Recent evidence suggest that after the initial outbreaks, the human-infecting flu A (H7N9) and local avian A/H9N2 strains had undergone further reassortment which may be responsible for the sudden increase in human cases in early 2014 [73]. It should be noted that HPAI A/H5N1 took about five years to attain the number of cases A/H7N9 is currently racking up. Such impressive disease burden catapults A/H7N9 alongside HPAI A/H5N1, if not ahead, as the next potential human pandemic contender. Unlike HPAI A/H5N1 viruses which cause massive die-offs in affected poultries, A/H7N9 and A/H10N8 do not induce noticeable clinical disease indicators. Such feature would make virus tracking and surveillance more difficult and challenging. In another light, the constant inclusion of A/H9N2 viral components into the zoonotic A/H5N1, A/H7N9, and A/H10N8 strains raises the question whether A/H9N2 viruses predominantly circulating in Chinese poultry could have adapted to support human infections when mixed with the right combination of segments (e.g., HA and NA).
Overall, these zoonotic transmissions validate the potential of AIVs to cause the next human pandemic. With the continued circulation of these viruses, the generation of more virulent and human-transmissible variants, however remote at the moment, remains a possibility given the right conditions and appropriate hosts. Hence, these concerns collectively call for continued virus monitoring and surveillance in humans and animal populations alike, worldwide. Coupled with virus characterization and pathobiologic assessment, these will help identify which strains we should be particularly worried about and set focus on in the future. Finally, the recent incursions of variant flu strains in humans, apart from H5 and H7 viruses, also require the need for appropriate virus-specific or efficacious universal vaccines that can induce broad cross-protective responses which can be made available and administered in the event of or even before a pandemic happens.

 XML Download
XML Download