

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Hydrocephalus is a common neurosurgical disease that develops in various conditions such as congenital anomaly, trauma, tumor, infection, etc. Ventriculoperitoneal (VP) shunt and endoscopic third ventriculostomy are the main methods of management. However, the high rate of complication and the limitation of indications of these surgical methods remain unsolved problems for the management of hydrocephalus.1212528) Much of the water pathophysiology of the ventricles as well as the brain parenchyma is still not elucidated. It is conceivable that further knowledge of brain water physiology, including cerebrospinal fluid (CSF) dynamics, may be a useful basis of management for related disorders.
Aquaporin (AQP) is a recently discovered protein that regulates fluid homeostasis in various organs, and several subtypes have been reported. Among the subtypes of AQPs, AQP 1 and 4 are abundant in the brain and are related to the regulation of water physiology.2029) AQP 1 is abundant in the choroid plexus where it regulates CSF physiology, and AQP 4 is distributed in the brain parenchyma.5101618)
Kaolin-induced hydrocephalus model is most commonly used one for the experimental research of the disease.815) Kaolin is one of the inert silicate and produces communicating and non-communicating hydrocephalus in accordance with the injection site by combination effect of physical deposition and local fibrotic response in the arachnoid and pial membranes.1231)
Author designed this experiment to study changes in the expression of AQP 1 and 4 in the kaolin-induced hydrocephalic model to elucidate water physiology in hydrocephalus and provide basic knowledge for the future development of treatment methods of hydrocephalus by regulating water physiology and protecting the brain from the disease.
Go to : 
Materials and Methods
This experiment was approved by the Institutional Review Board for Animal Experiments, and performed under the guidelines of the Institutional Animal Care Committee.
Animal model
Sprague-Dawley rats weighting 400 to 500 mg were used for the experiments. All experiments were performed on rats anesthetized with a mixture of ketamine (75 mg/kg), acepromazine (0.75 mg/kg), and xylazine (4 mg/kg). Each rat was immobilized in a home-made apparatus designed for easy cisternal injection with the neck flexed. Experimental hydrocephalus was induced by percutaneous injection of 0.06 mL of 12.5% kaolin solution mixed with 0.9% normal saline into the cisterna magna. The same volume of sterile saline was injected into the cisterna magna of control animals. Animals were sacrificed by injection with intraperitoneal ketamine on 3rd, 7th, 14th, and 30th days after injection. The brain was dissected to obtain the choroid plexus and frontal lobe parenchyma of each rat. Each experimental group was composed of 3 animals.
Western blot
Proteins were extracted from the choroid plexus and parenchyma, which were then dissolved in lysis buffer solution (25 mM Tris-Cl pH 7.6, 150 mM NaCl, 1% Nonidet P-40 [NP-40], 1% sodium deoxycholate, 0.1% sodium dodecyl sulfate [SDS], and protease inhibitor cocktail). The samples were centrifuged at 13,000 rpm for 30 minutes at 4℃. The supernatant was separated and the proteins were quantified by bicinchoninic acid assay (BCA) protein assay (Sigma, St. Louis, MO, USA). Protein samples were separated on 10% SDS-polyacrylamide electrophoresis gel and transferred to membranes (Millipore, Bedford, MA, USA). The membranes were blocked with 5% non-fat dry milk in Tris-buffered saline containing 0.1% Tween-20 for 1 hour at room temperature. The membranes were then incubated with anti-AQP 1,-AQP 4, or glyceraldehyde 3-phosphate dehydrogenase (GAPDH; Santa Cruz Biotechnology, Santa Cruz, CA, USA) at 4℃ overnight, followed by incubation with peroxidase-conjugated secondary antibody for 1 hour at room temperature. Signals were detected using an enhanced chemiluminescence (ECL) system (Amersham Biosciences, Buckinghamshire, England). Band densities were calculated using Image J, version 1.46r, computer-assisted image analyzer (National Institutes of Health, Bethesda, MD, USA).
Immunocytochemistry (IHC), and immunofluorescence (IF)
Rats were deeply anesthetized with an intra-peritoneal injection of ketamine and were perfused intracardially with 4% paraformaldehyde in 0.1 M sodium phosphate buffer (PB; pH=7.4). Then the tissues were dehydrated and paraffin embedded, and 4-mm coronal sections were taken from the brain. To detect AQP 1 and AQP 4, sections were deparaffinized with xylene and rehydrated in graded alcohol. Antigen retrieval was performed by autoclave heating in sodium citrate buffer. Endogenous peroxidase activity was quenched with 3% hydrogen peroxide and nonspecific binding was blocked with 4% bovine serum albumin. Samples were incubated overnight at 4℃ with a 1:200 dilution of anti-AQP 1/AQP 4 (Santa Cruz Biotechnology) and then rinsed with 0.1 M PB. This was followed by incubation in a 1:200 dilution of biotinylated anti-mouse IgG, and anti-rabbit IgG (Sigma, St. Louis, MO, USA) in 3% bovine serum albumin at 25℃ for 2 hours. The sections were then incubated in a 1:50 dilution of avidin-biotinylated horseradish peroxidase (Vector Laboratory, Burlingame, CA, USA) in 0.1 M PB for 2 hours and rinsed in 0.25 M Tris. Finally, staining was visualized by reaction with 3, 3′-diaminobenzidine tetrahydrochloride (DAB) and hydrogen peroxide in 0.25 M Tris for 10 minutes using a DAB reagent set (Kirkegaard & Perry, Gaithersburg, MD, USA). All sections were then rinsed in 0.1 M PB, mounted on Superfrost Plus slides (Fisher, Pittsburgh, PA, USA) and dried overnight at 37℃. The dried mounted sections were dehydrated with alcohol, cleared with xylene, and covered using coverslips with Permount mounting medium (Fisher). The labeled cells were identified and counted with attention to the separation of antibodies in three types of tissue of each animal. The labeled tissues were photographed using a Zeiss Axiopan microscope with high power differential interference contrast optics (Carl Zeiss, Oberkochen, Germany). For IF, brain sections were blocked with 4% bovine serum albumin for 1 hour at room temperature, incubated overnight at 4℃ with anti-AQP 1/AQP 4, then rinsed in 0.1 M PB. Slides were incubated in a 1:200 dilution of Alexa Fluor 594 goat anti-mouse IgG and Alexa Fluor 488 goat anti-rabbit IgG in 0.1 M PB containing 4% normal serum and 0.5% Triton X-100 at 25℃ for 2 hours. All sections were then rinsed in 0.1 M PB and mounted on Superfrost Plus slides (Fisher). The labeled tissues were photographed using an Axioplan-2 epifluorescence microscope (Carl Zeiss). Quantification of AQP 1 and 4 immunostaining was done using Labworks, version 4.5, computer-assisted image analyzer (UVP, Upland, CA, USA).
Statistical analysis
All statistical comparisons were computed using SPSS 17.0 for Windows (IBM Corp., Chicago, IL, USA). Data are expressed as mean±standard error of the mean. Repeated measure analysis of variance was used to compare groups. Null hypotheses of no difference were rejected if p-value were less than 0.05.
Go to :
Results
Hydrocephalic model
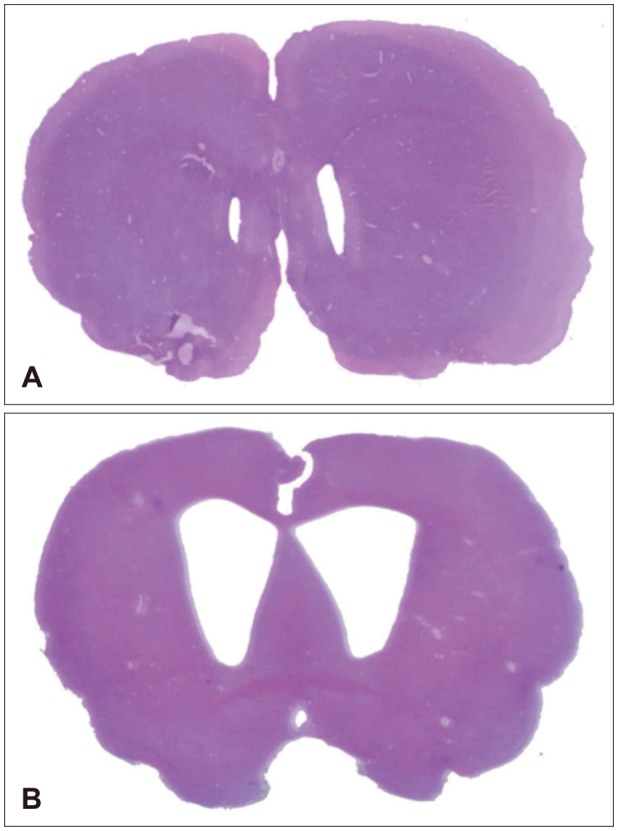
After intracisternal injection of kaolin suspension, progressive enlargement of ventricles was identified showing successful development of the animal model (Figure 1).
Changes in AQP 1 expression
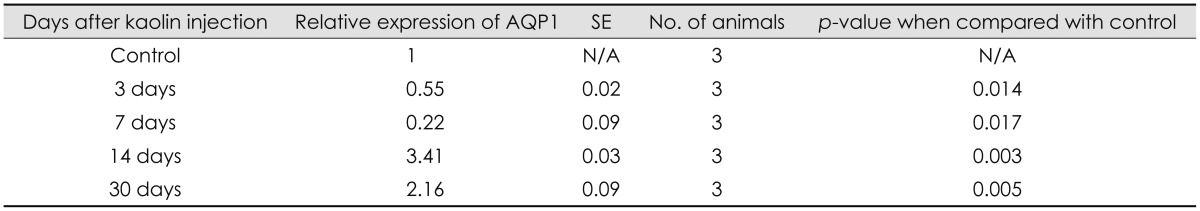
Expression of AQP 1 protein of the choroid plexus, measured by western blot demonstrated a significant decrease when measured at 3 and 7 days after kaolin injection, 55% and 22%, respectively, relative to control animal levels (p<0.05). The expression increased to 3.4 and 2.2 times higher than in the control at 14 and 30 days, respectively (p<0.05) (Table 1 and Figure 2). In other words, the chronological changes of APQ 1 expression showed decreased levels at the early stage and increased to levels, higher than the control, at the late stage (14 days), demonstrating a J-shaped curve.
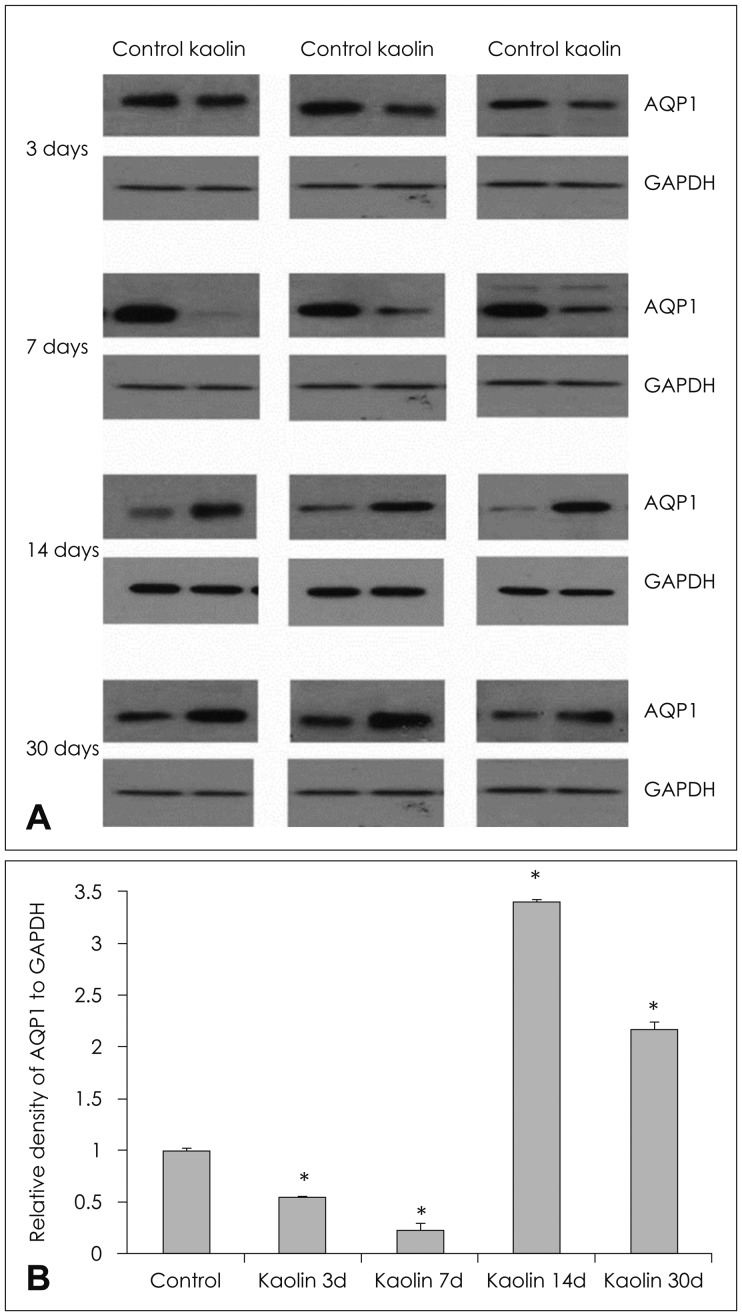 | FIGURE 2(A) Expression of aquaporin (AQP) 1 in the choroid plexus of hydrocephalic and control rats in western blot analysis of each experimental animal and (B) changes according to time interval after intracisternal kaolin injection. Expression decreased progressively on day 3 and day 7, but was higher than that of the control at day 14. (n=3, *p<0.05; for detailed p-value; Table 1). GAPDH: glyceraldehyde-3-phosphate dehydrogenase.
|
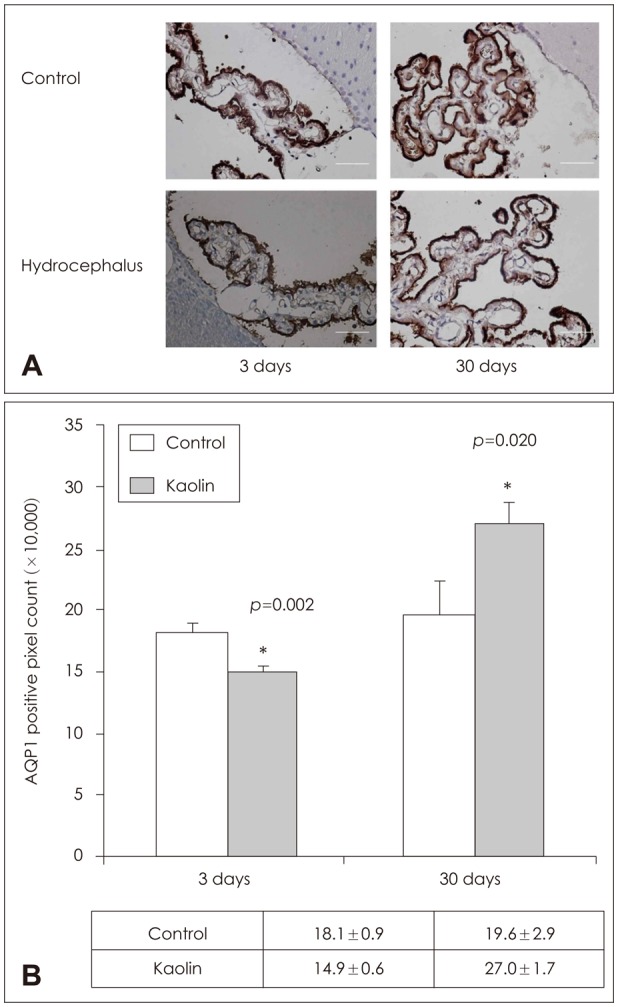
IHC staining showed a statistically significant increase in number of AQP 1 staining epithelial cells in the choroid plexus at the positive pixel count, corresponding with the findings of the findings of western blots (Figure 3).
Changes in AQP 4 expression
Parenchymal AQP 4 measured by western blot did not change significantly at 3 days after kaolin injection. Expression decreased at 7 days and increased at 14 days, 62% and 275%, respectively, compare to the control (p<0.05) (Table 2 and Figure 4). It remained high level at 30 days after injection. IHC revealed that the number of AQP 4 positive pixel increased in both 3 and 30 days rats (Figure 5). AQP 4 expression measured by western blot at 3 days was not statistically significant. However, it was noted to be significantly higher in positive pixel count in IHC. Changes in AQP 4 expression in early stage hydrocephalus were variable and require further examination. IF staining showed that AQP 1 was expressed in choroid plexus epithelium but not in the parenchyma at both 3 and 30 days. AQP 4 was expressed in the brain parenchyma, while it was not expressed in the choroid plexus (Figure 6).
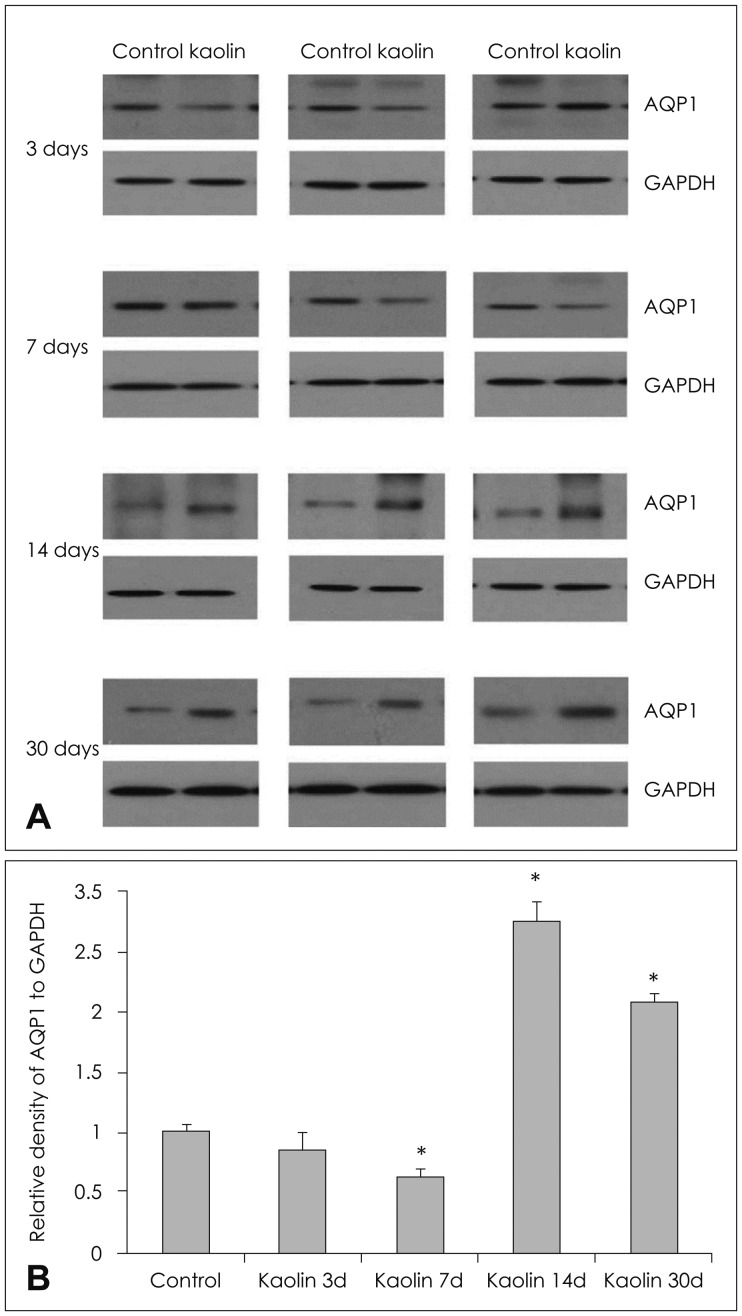 | FIGURE 4(A) Expression of aquaporin (AQP) 4 in the brain parenchyma of hydrocephalic and control rats in western blot analysis of each experimental animal and (B) changes according to time interval after intracisternal kaolin injection. Expression decreased early after injection, but increased to higher levels than that of the control after 14 days (n=3, *p<0.05; for detailed p-value; Table 1). GAPDH: glyceraldehyde-3-phosphate dehydrogenase.
|
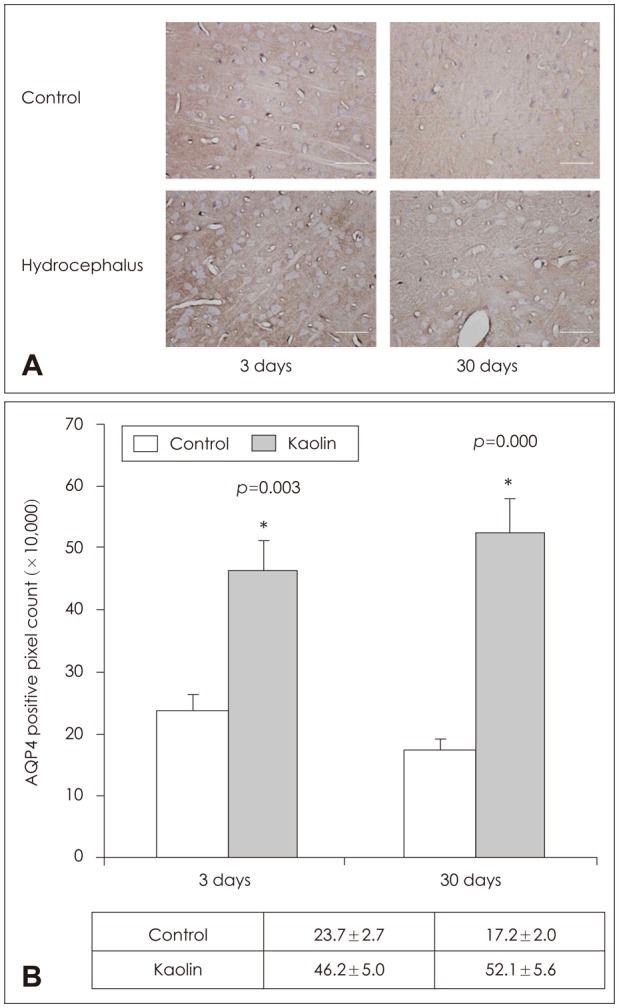 | FIGURE 5Aquaporin (AQP) 4 immunolabeling in the parenchyme. (A) Immunohistochemical staining and (B) quantificative measurement by positive pixel count of AQP4 of the brain parenchyma revealed expression to be higher than in the control at day 3 and day 30 (n=3, *p<0.05).
|
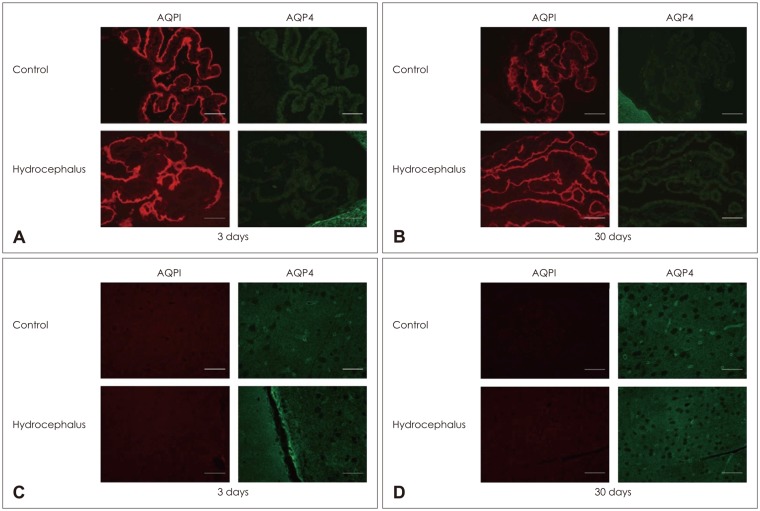 | FIGURE 6(A, B) Immunofluorescence staining of aquaporin (AQP) 1 and AQP 4 in the choroid plexus and brain parenchyma in hydrocephalic rats after 3 days and 30 days. AQP 1 was expressed in the choroid plexus both in control and in hydrocephalic rats, but was not expressed in the brain parenchyma. (C, D) In contrast, AQP 4 was expressed in the brain parenchyma, but not in the choroid plexus.
|
Go to :
Discussion
Hydrocephalus is a relatively common neurosurgical disease occurring congenitally or in association with many other diseases. VP shunt is a primary treatment method for the disease. In spite of the many advancements of the shunt system, the complication rate is still high and the revision rate reaches approximately 30%.1212528) Endoscopic third ventriculostomy is another good method for the management of hydrocephalus. With this method, the patient is free from the possible complications of VP shunt. However, the relatively high failure rate in communicating hydrocephalus type is still a limitation of endoscopic third ventriculostomy.49) No medical treatment provides a prolonged effect. Acetazolamide and furosemide reduce the production rate of CSF and is prescribed for short-term durations.632) They are effective in transiently reducing the intracranial hypertension associated with hydrocephalus; however, they are not applicable for the long-term treatment of hydrocephalus. Knowledge of brain water physiology is expected to provide the basis of future medical management of hydrocephalus.
AQPs are membrane transport proteins that were initially identified in red blood cells and kidneys by Preston and Agre24) in 1991 as proteins regulating osmotic water transport, now known as AQP 1. Thereafter, many subtypes were identified and found to be related to various diseases in humans. AQP 1 and 4 are abundant in the brain. AQP 4 is located abundantly in the brain parenchyma, particularly in astrocyte endfeet lining the blood-brain barrier and glia limitans, and is related to brain water physiology, including brain edema.1629) It is known that abnormalities in AQP 4 are related to many disease states. Neuromyelitis optica is well known as a disease caused by AQP 4 abnormality.1118) AQPs are also related with brain edema, cancer cell migration, and epilepsy.18) Modification of AQP has been proposed as an adjuvant treatment of these diseases,2030) even though the progression is very slow.
AQP 4 functions in the physiology of water movement in the brain. It facilitates water accumulation in cytotoxic edema and the clearance of excess brain water in vasogenic and interstitial edema.2930) Upregulation of AQP 4 has been found in various disease states, such as trauma, stroke, and hydrocephalus.142226) In acute hydrocephalus, the brain parenchyma surrounding the ventricle is a subject of interstitial edema.1920) AQP 4 is upregulated in association with excess water content in the interstitial space as a compensatory response to facilitate CSF elimination.20) In the kaolin-induced hydrocephalus model in AQP 4 deficient mice, the progression of hydrocephalus was accelerated.3) The excess fluid in the interstitial space can become clear as a result of AQP 4 in the subarachnoid space or blood.20)
AQP 1 is mainly distributed in choroid plexus ependyma facing the ventricle in the brain where it is involved in CSF secretion. Oshio et al.17) reported that deletion of AQP 1 in mice reduced intracranial pressure by a slowing of CSF secretion. In acute hydrocephalus, blockage of ventricular circulation induces a reduction of CSF production by at least partially reducing AQP 1 expression. Paul et al.22) found biphasic expression of AQP 1 in the congenital hydrocephalus model, representing the compensatory mechanism of choroidal CSF secretion. The reduced AQP 1 level early after birth becomes elevated to normal levels in delayed hydrocephalus. The levels of AQP 1 protein expression and AQP 1 mRNA were increased in the cortex and pia. In contrast, AQP 1 expression was decreased in the ependyma of late hydrocephalus. According to researchers, immunostaining revealed the AQP 4 protein to be increased in astrocyte endfeet, ependymal, and pia.22) In advanced stage hydrocephalus, clearance of excess parenchymal fluid from ventricles is made through increased AQP 4 expression. In the late stage, AQP expression returned to levels seen in the control in the choroid plexus; however, it was higher in the ependyma, pia, and subependyma. In our experiment, AQP 1 levels in the late stage were significantly higher than in control animals. The mechanism of the increase in AQP 1 expression in the late stage of hydrocephalus is not clear. Paul et al.22) hypothesized upregulation due to pressure-recruited neuroendocrine modulators. Atrial natriuretic peptide (ANP) is a regulator of AQP 1 channel function.7) ANP down-regulates CSF formation via neuroendocrine effects on choroid plexus epithelium.722) However, this overexpression should be confirmed with further studies. Moreover, changes at later stages of chronic hydrocephalus should be clarified.
Our research was performed in acquired hydrocephalus rats that had had normal brain development before hydrocephalus, in contrast to the congenital model of Paul et al.22) where congenital anomalies of AQP expression could not be excluded. In our experiments, choroidal AQP 1 was progressively decreased at 3 and 7 days after kaolin injection and increased at 30 days, an interesting finding, but the mechanism and its effect on hydrocephalus are not clear and require further investigation.
Parenchymal AQP 4 changes were parallel to AQP 1 changes except in the early stage. Regarding increased expression of AQP 4 30 days after injection in our experiment, Mao et al.14) also reported that in their kaolin induced rat model, mRNA expression was higher at 3 weeks than in controls, and this expression remained at 9 months after injection. They explained this effect as a compensatory mechanism of hydrocephalus. This finding corresponded with our experimental rats at 30 days after injection of kaolin. However, in their report, the protein level examined by western blot was not elevated. They explained this discrepancy as occurring due to redistribution and increased turnover of the protein, though this requires further confirmation. Shen et al.26) reported increased AQP 4 expression in their arrested congenital hydrocephalus model and related this with the development of an alternative pathway of CSF circulation. According to them, AQP 4 is abundantly distributed in endfeet of astrocytes and glial limiting membrane in the subpia and subependyma as well as the perivascular space. These structures could be roots of the compensatory alternative CSF pathway in hydrocephalus.
It is conceivable that compensation for acute hydrocephalus, mostly or at least partly, occurs with decreased CSF production, while in later stages, the parenchymal fluid clearance mechanism predominates. However, confirmation with further research is still needed to explain the greater expression of AQP 1 and 4 in the choroid plexus and brain parenchyma in later stages. Also, more pathophysiological information about CSF physiology is needed in the young age model because many of our hydrocephalus patients are children with developing brains.
The upregulation of AQP 4 or downregulation of AQP 1 in the brain can provide us with important knowledge for the development of alternative regimens for the management of hydrocephalus or for the protection of the brain from various disease states. Even though AQPs are expected to provide important targets for the management of diseases related to water physiology, including hydrocephalus, reports on the expression of AQPs in hydrocephalus are heterogenous both in humans and in animals.2510) Expression of AQP 1 in human hydrocephalus is seldom reported, and results are heterogenous.1327) It is mainly reported in patients with choroid plexus lesions.1327) Moreover, the CSF outflow system of humans differs greatly from that of animals. In animal, especially low animals, the lymphatic system plays a more important role than the arachnoid system,23) while in humans, the role of the lymphatic system is not clearly defined. We require further information about the physiological mechanism related to AQPs and water physiology.10)
A limitation of our research was that the hydrocephalus model was verified at the beginning of the experiment in animals for model verification. For the better credibility of the experiment, each experimental rat should be confirmed by radiographic examination such as computed tomography scan or magnetic resonance imaging before sacrifice. However, it was not possible in our laboratory to confirm the hydrocephalus in living animal by radiography.
Go to :
Conclusion
The changes in expression of AQP 1 and 4 in experimental hydrocephalic rats induced by intracisternal injection of kaolin were evaluated. The expression of AQP 1 decreased sharply in the early stage of hydrocephalus and suggested compensatory reduced production of CSF. Expression was elevated to higher levels than those of controls in the later stage. This is hypothesized to be related to other mechanisms of CSF secretion such as neuroendocrine function.22) AQP 4 was variable in the early stage but higher in the later stage, suggesting a compensatory mechanism for the clearance of excess interstitial fluid in hydrocephalus. We require more information about brain water physiology to develop alternative treatment regimens for the management of hydrocephalus and the protection of the brain from damage.
Go to :
 XML Download
XML Download