

 PDF
PDF ePub
ePub Citation
Citation Print
Print


C57BL/6 mice—the multipurpose model inbred strain—are average-sized laboratory mice most commonly used in various research fields [1]. High reproductivity of C57BL/6 and their substrains [2] has made them especially useful in reproduction studies. Blastocytes originating from the C57BL/6 strain are easy to handle and can be used in gene transformations of varying degrees as well as in the production of knock-out and knock-in mice. Among the diverse and well-established substrains, C57BL/6J and C57BL/6N are currently being employed by breeding centers, industrial organizations, and research institutions across the globe [3]. Because they have been widely used for a long period, a large amount of historical background data has been accumulated in the fields of histopathology, hematology, blood biochemistry, and urinalysis/urine biochemistry. These historical data are essential for interpreting results and improving the reliability of animal experiments. However, in the field of histopathology, spontaneous data, especially on non-neoplastic lesions, for relatively younger mice that are more frequently used in general research are not yet abundant, in contrast to the data on neoplastic lesions for older mice aged one year or more. Various types of non-neoplastic lesions can also occur in healthy animals, and their incidence and severity can be increased by certain test items and experimental conditions [4]. Thus, for more accurate histopathological analyses, spontaneous changes should be determined precisely, and the pathologist should be well aware of the different spontaneous changes that occur according to the species and age of the lab animal. For histopathological determination of disease or toxic changes, the accumulation of data is essential, because the basic data for spontaneous histopathological lesions allow an appropriate examination of the incidence and severity of the condition and provide a crucial standard for accurate determination of the changes caused by test items or conditions [4]. For more reliable toxicological assessment and in vivo research, the present study performed toxicological pathology analyses of the major organs (liver, spleen, kidney, thymus, heart, testis, epididymis) of 10- and 13-week-old C57BL/6N mice, which are frequently used in general short-term tests.
Materials and Methods
Animal experiment design
For the experiment regarding the effects on the animal supplying facilities, 60 male mice of the C57BL/6N strain were procured from each of two institutions: A (from Korea) and B (from Japan). The mice were six weeks old upon collection, and permitted an acclimation period of 5 days to the laboratory environment before being separated into 10-week-old (G1, G3) and 13-week-old (G2, G4) autopsy groups, each containing 30 mice. The studies involving groups G1 and G2 were performed one year prior to those involving G3 and G4. The design of the experimental groups is given in Table 1.
Each mouse was bred in a stainless steel breeding cage, under the general experimental conditions of the Good Laboratory Practice (GLP) animal facility. Given that the purpose of the experiment was the accumulation of historical data, the mice were provided ad libitum food and drinking water without any handling procedures such as injections until the terminal sacrifice day.
All animal experiments in this study were carried out according to the standard operating procedures (SOPs) approved by the Animal Protection Act of Republic of Korea and the Guide for the Care and Use of Laboratory Animals (by ILAR publication), after the acquisition of IACUC approval and the relevant study numbers (Study No. B215032, JB14049).
Terminal procedure and histopathological analysis
All mice used in the experiment were fasted for 4 hours on their corresponding final sacrifice day (the day when the mice became approximately 10 or 13 weeks old). Isoflurane was used for the deep sedation and animals were sacrificed by exsanguination from the vena cava and aorta, which was followed by external examination of the abdominal, thoracic and cranial cavities. From each sacrificed mouse, the weights of the major organs—liver, kidney, heart, thymus, spleen, testis, epididymis—were measured, after which each organ was fixed. The testis and epididymis from each mouse were fixed in Bouin's fixative for 48 hours and transferred to a solution of 70% ethanol. The rest of the organs were fixed in 10% normal buffered formalin. The fixed tissues of the organs were routinely processed to slides, stained with hematoxylin and eosin (H&E), and examined microscopically.
Historical data analysis affiliated institution
Among the experiments carried out in the affiliated research institution, the data from the histopathological tests of the epididymides were analyzed, and the results were printed out and gathered using the path/tox system (Version 4.2.2, Xybion Medical system Co., Lawrenceville, NJ, USA) and Pristima (Version 6.4.0, Xybion Medical System Co., Lawrenceville, NJ, USA). Among the experiments carried out from 2014 until 2017, the data from the histopathological tests on C57BL/6N, C3H/Hen, Balb/cAnN, and ICR mice and Sprague-Dawley (SD) rats were analyzed. Before the analysis, the data were classified into two categories: animals that were 30 weeks old or younger and those that were older than 30 weeks.
Immunohistochemical (IHC) staining
After the H&E microscopic inspection, the epididymides were used for serial sectioning and subsequent immunohistochemical staining, to histopathologically assess the mechanism underlying the formation of sperm granulomas caused by tubule injury. The tissue samples were stained using the following antibodies: anti-high mobility group box 1 (HMGB1, Thermofisher Scientific, Waltham, MA, USA), anti-activated cysteine-dependent aspartate-directed proteases-3 (activated caspase-3, Abcam plc, Cambridge, UK), anti-cleaved-poly(ADP-ribose) polymerase (c-PARP, Abcam plc, Cambridge, UK), anti-microtubule-associated protein 1 light chain 3 beta (LC3B, Abcam plc, Cambridge, UK), and anti-superoxide dismutase 2 (SOD2, GeneTex, Irvine, CA, USA). The 1st antibody sources and concentrations were described in Table 2. Immunohistochemical staining was carried out using automated immunohistochemistry slide staining system (Discovery XT, Ventana Medical System, Tucson, AZ, USA). The antigen restoration for caspase-3 involved the citrate buffer and Tris buffer for HMGB1, c-PARP, LC3B, and SOD2. All primary antibodies were allowed to react for three hours, before the 16-minute secondary reaction with the universal secondary antibody (RUO, Ventana Medical System, Tucson, AZ, USA). 3,3-Diaminobenzidine (DAB) was used for color development before the microscopic examination of the antibodies under a light microscope (Axioplan2, Zeiss, Oberkochen, Germany). For the comparison with C57BL/6N mice, the same procedures were used for staining the epididymides from ICR and BALB/c nude mice and SD rats of the similar age.

Table 2
Antibody sources and concentrations
![]()
Statistical analysis
The data were analyzed for homogeneity of variance, using a folded f-test. Homogeneous data were analyzed using Student's t-test, and heterogeneous data were analyzed using Welch's t-test. Statistical analyses were performed using SAS 9.4 (SAS Institute, Cary, NC, USA). All tests were two tailed and P<0.05 was considered significant.
Go to : 
Results
Spontaneous lesion in epididymis of C57BL/6N
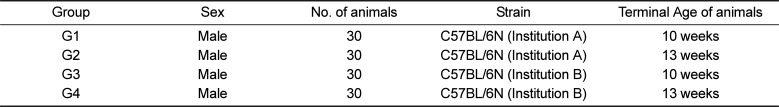
In the macroscopic observation during the necropsy, nodules in epididymis and enlarged epididymis were observed. In G1, there were three cases of enlarged epididymis; G2, nine cases; G4, two cases. The nodules were observed in a two cases in G1. The histopathological tests on the collected and fixed organs showed mononuclear cell infiltration and focal necrosis in the liver; tubular basophilia, tubular dilatation, mononuclear cell infiltration, pyelonephritis, and hyaline casts in the kidney; mixed cell infiltration in the heart; and extramedullary hemopoiesis and lymphoid hyperplasia in the spleen. These observations were of the same level of incidence and severity as the general spontaneous changes that occur in other mice (ICR) of similar ages. However, what attracted our attention was a unique non-neoplastic spontaneous lesion, known as a sperm granuloma, found in the epididymis of all groups, displaying higher incidence and severity in our test species than in the other lab animals (Figures 1, 2). The nodules and enlarged epididymis in macroscopic observation corresponded to various forms of sperm granulomas, which were also detected in the healthy epididymis upon macroscopic observation. The sperm granulomas observed in the epididymis were mostly detected unilaterally. In the case of animals in G1 and G2 (from Korea), the lesion showed higher incidence in 10 weeks autopsy animals than in 13 weeks autopsy animals. In the case of animals in G3 and G4 (from Japan), relatively lower incidence was observed in 10 weeks autopsy animals, showing an increasing trend in 13 weeks autopsy animals. The changes detected in the epididymis are summarized in Table 3.
 | Figure 1Tail of the epididymis of 10-week-old C57BL/6N mice (G1 group). Sperm granulomas (arrows) are shown in the tail of the epididymis on the right, in contrast to the tail of a healthy epididymis on the left (a). Bar size; 2 mm. High magnification image showing a sperm granuloma in of 10-week-old mice; the sperms (S) leaking out of the tubules are visible at the center, together with granulomatous inflammation in the surrounding tissues. Sperms discharged into the interstitial space are being actively devoured by macrophages (arrowheads) (b). Bar size; 100 µm. High magnification image showing the epididymis body; the damaged tubule (arrows) from which the sperms leaked is indicted (c). Bar size; 200 µm.
|
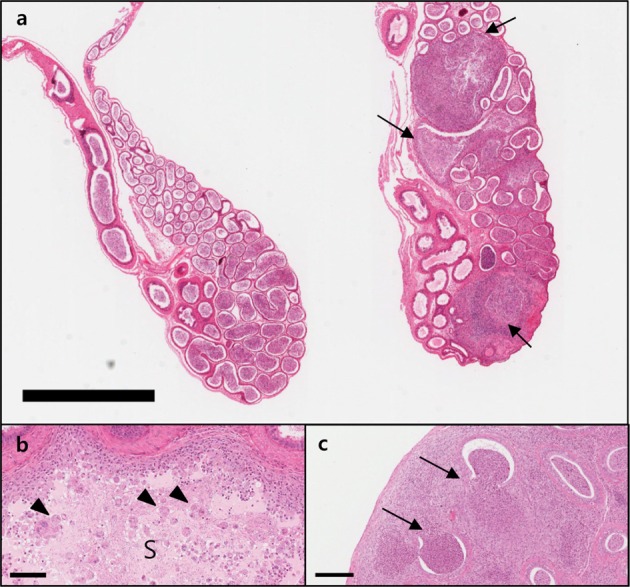
 | Figure 2High magnification image showing the changes associated with sperm granuloma in the epididymis of 10-week-old mice (G1 group). Absence of sperm in the lumen (L, oligospermia) and hyperplasia of clear cell (arrows) in epithelial cells (a). Macrovacuolation (arrowheads) in cytoplasm of epithelial cells (b). Bar size; 200 µm.
|

The observed sperm granulomas had the shape of a typical granuloma. The sperm core is found in the center of the interstitial space next to the tubules and is surrounded by chronic inflammatory cells, such as macrophages and lymphocytes, and fibrous granulation tissues. In the epididymides with sperm granulomas, tubules in tail of epididymis displayed oligospermia (absence or reduction of sperm in the epididymal tubules) and clear cell hyperplasia. Oligospermia was observed as a tubule with an empty space around the granuloma. Clear cell hyperplasia was visualized as increased number and size of clear cells in tubular epithelium with oligospermia. In all tissues where sperm granulomas were observed, oligospermia/clear cell hyperplasia, cellular debris, and tubular vacuolation were consistently observed. Moreover, judging by the pattern of incidence, oligospermia/clear cell hyperplasia, cellular debris, and tubular vacuolation not accompanying a sperm granuloma may indicate that the condition might exist in the parts of the epididymis not shown on the slides. The forms of the detected changes were similar across the animals from A and B institutions.
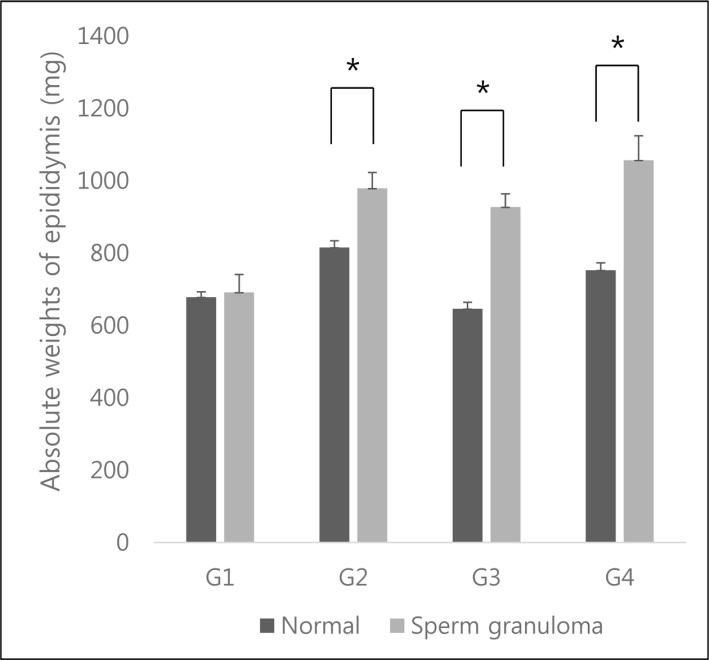
In the epididymides of increased weight, histopathological analysis revealed them to contain sperm granulomas, as opposed to healthy epididymides. The significantly increased organ weight for 10-week-old autopsy animals from Institution A (G1) and 13-week-old autopsy animals from Institution A and B (G2, G4) are shown in Figure 3.
Historical data analysis
Historical data analysis was performed to compare the sperm granulomas observed in C57BL/6N mice with the incidence of other laboratory animals. The results showed approximately 1–1.4% incidence in ICR and SD rats. In the case of C57BL/6N mice, 5% incidence was reported in animals older than 30 weeks. The results are summarized in Table 4.
Immunohistochemistry in epididymides of C57BL/6N
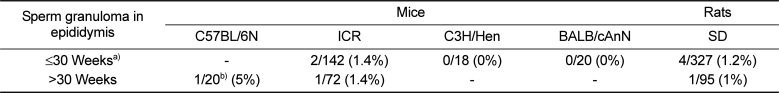
The results of the immunohistochemical staining of C57BL/6N mice epididymis, involving the anti-high mobility group box 1 (HMGB1), anti-cysteine-dependent aspartate-directed proteases-3 (caspase-3, active form), anti-microtubule-associated protein 1 light chain 3 beta (LC3B), and anti-superoxide dismutase 2 (SOD2) antibodies, showed no significant chromatic figure from the HMGB1 and LC3B antibodies, whereas the caspase-3 and SOD2 antibodies led to significant chromatic figures. In the case of caspase-3, Immunohistochemistry was performed in 12 healthy epididymides and 8 epididymides with sperm granuloma (A total of 20 epididymides from 10 mice) and signals were detected in single cell units on the epithelial cells of the epididymis tubules in all samples, regardless of the presence of sperm granulomas; however, 2 cases of distinctly increased caspase-3 chromatic figures were observed only in epididymides with sperm granuloma. In the case of SOD2, the chromatic figure was found in interstitial cells and in the internal cilia of the tubules located in the tail of the epididymis. Regarding the two antibodies that produced significant chromatic figures, counterstaining was carried out using epididymides from ICR and BALB/c nude mice and SD rats of similar ages. In the case of caspase-3, the significantly increased chromatic figures observed in C57BL6/N mice were not observed in the other species of lab animals (Figure 4). In the case of SOD2, the various species of animals displayed similar patterns, with only a minor degree of difference (Figure 5). The results of the immunohistochemical staining involving caspase-3 in the four different lab animals are described in Table 5.
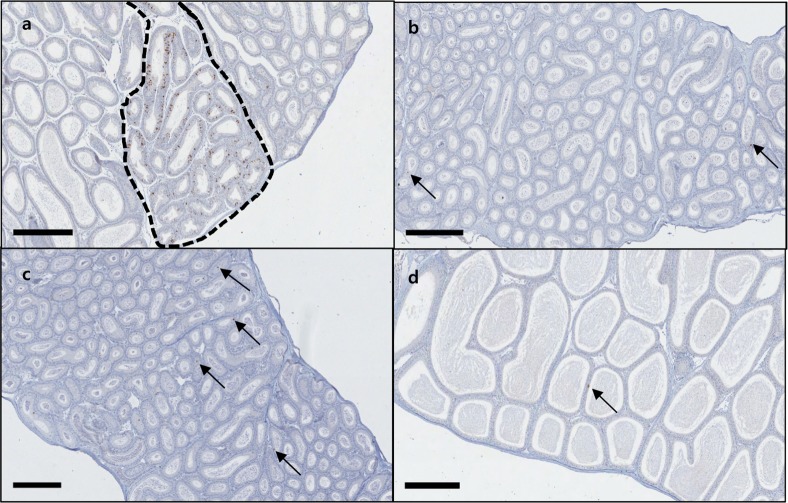 | Figure 4Image of the caspase-3 IHC staining in epididymis of lab animals. Significantly more caspase-3 chromatic figures are observed in the tubules of the C57BL/6N mouse epididymis (dotted line) than the other lab animal species, in which 1-2 chromatic figures are observed per tubule (arrows). C57BL/6N mouse (a), ICR mouse (b), BALB/c nude mouse (c), SD rat (d). Bar size; 400 µm.
|
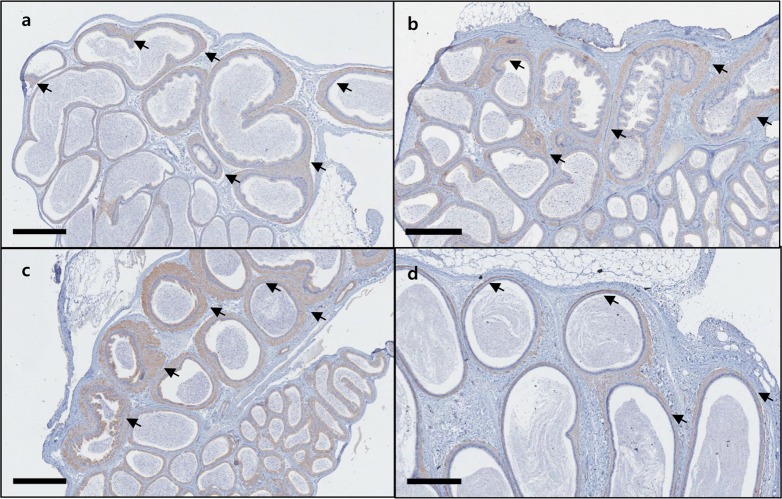 | Figure 5Image of the SOD2 IHC staining in epididymis of lab animals. Positive SOD2 chromatic figures were observed in the tubules in tail of epididymis. C57BL/6N mouse (arrows) (a), ICR mouse (b), BALB/c nude mouse (c), SD rat (d). The chromatic figure was found in interstitial cells and in the internal cilia of the tubules. Bar size; 400 µm.
|

The epididymis tissues that exhibited caspase-3 chromatic figures were serially sectioned and additionally stained with anti-cleaved-poly(ADP-ribose) polymerase (c-PARP). The results showed significant chromatic figures of c-PARP in the same tubules that exhibited the caspase-3 figures (Figure 6).
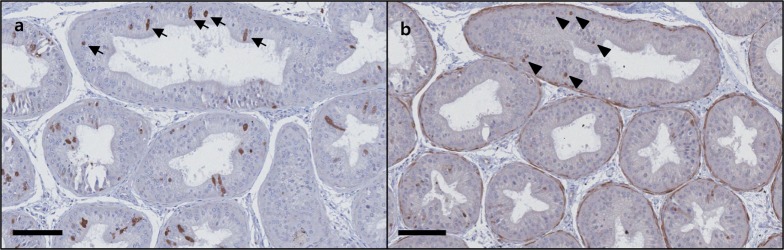 | Figure 6Results of c-PARP IHC staining for the area of the epididymis of C57BL/6N mice where the caspase-3 chromatic figure is found; caspase-3 (a), c-PARP (b). The staining of the serially sectioned area shows c-PARP chromatic figures (arrowheads) in the cellular nucleus of the tubules that exhibited caspase-3 chromatic figures (arrows). Bar size; 100 µm.
|
Go to :
Discussion
For the C57BL/6N mice procured from Korean and Japanese institutions that specialize in lab animals, nodules in epididymidis and enlarged epididymis were macroscopically detected. The pathological assessment detected significantly more frequent sperm granuloma incidence than that generally observed in other laboratory animals. In previous literature, few or no articles reported on sperm granulomas occurring in the epididymis of relatively young C57BL/6N mice (10 and 13 weeks old). In a study that presents historical data on the aging mice of inbred strains (129S1/SvW, A.CA-H2f/W, AKR/W, BALB/cW, BN/aW, C57BL/6 W, C57BL/10 W, C3Hwad/W, C3Hwad/W, CBA/W, DBA/2W, WOM/W and B6; 129), sperm granuloma did not occurred [56]. In CByB6F1-Tg(HRAS)2Jic mice, which are often employed as animal models for studying the carcinogenicity replacement, approximately 0~8% of sperm granuloma was reported [7]. Control historical data on RCCHan: wist rats report approximately 8.3% incidence in 4-week-old rats and 3.4% in 13-week-old rats [8]. In the case of Fischer344 rats, which are widely used in toxicology research, 0% of sperm granulomas occurred in 90-day-old animals (approximately 13 weeks) [9]. To compare the incidence of sperm granuloma observed in C57BL/6N mice with that observed in other strains, the historical data of the institutions in the present study were analyzed. The collection of data was restricted to the vehicle control group animals of recently performed studies (2014–2017). The animals were classified into two categories: those 30 weeks old or younger and those older than 30 weeks before the final sacrifice day. Analysis of data from other laboratory animals revealed lower incidences of sperm granulomas than observed in this study. Also, a significantly higher incidence of sperm granuloma was observed in C57BL/6N mice older than 30 weeks. The results showed approximately 1–1.4% incidence in ICR mice and SD rats. In the C3H/Hen and BALB/cAnN, sperm granulomas were not observed. In the case of C57BL/6N mice, the data were insufficient for direct comparison; however, 5% incidence was reported in animals older than 30 weeks. The incidence of sperm granulomas was lower in C57BL/6N mice over 30 weeks than in 10 and 13 weeks of age, suggesting that C57BL/6N mice are characterized by younger age. The factors causing granulomas are thought to decrease with age.
The testis and epididymis are closely related, and a lesion found in one can affect the other. Thus, when performing a histopathological assessment, the two organs must be tested in close proximity [1011]. The present study detected tubular dilatation (2/30)—a phenomenon correlated to epididymal sperm granulomas—in the testis of mice in G1 (Institution A, 10-week-old mice). However, the lesion was not observed in the testis of any other animal carrying epididymal sperm granulomas. In the testis of mice in G3 (Institution B, 10-week-old mice), tubular atrophy was detected in the testis (2/30); however, the condition did not accompany epididymal sperm granulomas. In addition, inflammation (chronic or chronic active form) was detected as a type of background lesion commonly observed in the preputial glands of rodents, without correlation to epididymal sperm granulomas. Sperm stasis, spermatocele, and tubular dilatation, the phenomena that characterize the rupture caused by epididymal tubular occlusion, were not detected in epididymis.
Sperm granulomas are granulomatous inflammation lesions that result from intense immune responses to sperms that have leaked out of the testis or epididymis due to a specific reason and become recognized as an auto-antigen. The cause of the damage to the strong defense walls of the testis and epididymis tubules is yet unclear; however, physical trauma or toxic substances are the presumed causes [12]. Occlusion of the vas deferens via experimental vasectomy is known to be correlated with nearly 100% incidence of sperm granuloma [13]. In addition to physical trauma, the lesion can be induced by the injection of chemical substances, where it occurs in specific parts (head, body, tail) of the organ [14].
Because the damage of the epithelial cell is a major cause of sperm granuloma, immunohistochemical staining was carried out to investigate the mechanism of cell damage. The four most frequently occurring cellular damages (HMGB1, caspase 3, SOD2, LC3B) were investigated. HMGB1 is a chromatin-associated protein of the non-histone group, which induces the proinflammatory response when it is secreted by cells involved in inflammation [15]. Caspase 3 is an important protein that belongs to the cysteine proteases, which play a central role in apoptosis [16]. LC3B is a protein found in a wide array of mammalian tissues, where it induces autophagic cell death [17]. SOD2 plays a crucial role in protecting the cells by converting superoxide into hydrogen peroxide and diatomic oxygen, when oxidative stress increases its secretion [18].
The results showed significant increase in the chromatic figures from caspase-3 and SOD2 antibodies in the epididymis of C57BL/6N mice. To determine whether the observed chromatic figures were specific to C57BL/6N mice, the same procedures were carried out on the epididymis of ICR and BALB/c nude mice and SD rats. The characteristic chromatic figure of caspase-3 was not detected in any other species of lab animals. The chromatic figure of SOD2 was detected in a similar pattern in the other animals. The chromatic figure of caspase-3 observed in the epididymis of C57BL/6N mice can be regarded as evidence for the existence of a signaling pathway in the apoptosis of epididymal tubular cell. The negative chromatic figures of other antibodies indicate that the proinflammatory response, autophagic cell death, and oxidative stress, which occur at the stage where the each target proteins are involved, are not likely to be correlated with the naturally occurring epididymal sperm granuloma of C57BL/6N mice.
To better understand the caspase-3-induced apoptosis, the area where the caspase-3 chromatic figures were observed was immunohistochemical stained with anti-c-PARP. The c-PARP is a protein that interacts directly with DNA to induce apoptosis when it is activated by caspase-3 [19]. It is, hence, an important marker of apoptosis. The results of immunostaining showed that intracellular caspase-3 is activated spontaneously in the epididymal epithelium and it is likely to be the factor that induces apoptosis by converting PARP into c-PARP and causing DNA fragmentation in nucleosomes. It is also likely that the damaged tubules lost the ability to separate the stroma from the sperms in the epididymis, allowing the sperms to be discharged and granulomas to occur.
Apoptosis under normal circumstances is an essential mechanism for the survival and development of organisms, such as the formation and regulation of tissue structures, homeostatic maintenance of cell numbers, and removal of abnormal or dangerous cells [20]. However, when apoptosis is induced abnormally, the result is a variety of diseases and lasting damages to the nerve cells leading to Huntington's disease, Alzheimer's disease, Parkinson's disease, amyotrophic lateral sclerosis, retinitis pigmentosa, or spinal muscular atrophy [2122]. Apoptosis is also known to be correlated with autoimmune diseases such as rheumatic arthritis [2324], atherosclerosis, and cardiomyopathy. In particular, a study reported an increase in caspase-3 expression in epididymal tubules after the injection of epichlorohydrin—an antifertility agent that is directly toxic to the epididymis and inhibits sperm motility [25]. The expression of caspase-3 in the epididymal tubules of C57BL/6N mice, detected in the present study, was not observed in ICR or BALB/c nude mice or SD rats, which implies that it is characteristic to C57BL/6N mice.
In the present study, the cause of the granulomas cannot be chemically based because there was no injection of chemical substances. The probability that the cause is due to physical trauma is also low, because the animals employed were for the accumulation of historical data and there was no physical handling involving any kind of injection. Moreover, each animal was bred in a single cage, which eliminates the probability of physical trauma caused by interaction with other animals. The laboratory where the study was carried out was an official GLP institution, which performed all procedures according to SOPs; hence, any factors related to external infection, feed, or laboratory conditions can be eliminated. In an experiment targeting other mice (ICR mice), carried out by the same investigators, using the same equipment, and in the same laboratory, the characteristic sperm granuloma was not observed. Moreover, given that the condition was observed consistently across the animals from two institutions—one domestic and the other overseas—the changes cannot be due to differences between breeding facilities. The higher incidence of sperm granuloma is considered strain-specific to C57BL/6N mice.
In the case of mice, as in the case of other lab animals, many strain-specific spontaneous diseases are known to occur. C57BL/6N—an inbred strain—also exhibits diverse and characteristic spontaneous changes [2627]. Several eye abnormalities such as microphthalmia and anophthalmia have been observed [28], and corneal opacities (corneal mineralization) occurs with approximately 5% incidence [29]. In addition, disruption and loss of outer and inner hair cells are observed in their cochlea, and collapse and degeneration are observed in their organ of Corti [3031]. A higher incidence of noise-induced hearing loss has been observed when compared with other mouse strains [3233].
To conclude, the present study has confirmed that spontaneous sperm granulomas can occur in 10- and 13-week-old C57BL/6N mice. Visual inspection could discriminate the more-severe cases of sperm granulomas, and a significant increase in absolute organ weight was observed in the epididymides with the granulomas, in comparison to that of healthy epididymides. The sperm granuloma detected in the epididymis is likely to affect the maturation and ejaculation of sperms, and the lasting chronic inflammation response may impart a systemic influence to the mice. Thus, when interpreting the results of an in vivo study employing C57BL/6N mice, especially when the study involves toxicological studies, a thorough consideration of epididymal sperm granulomas should be guaranteed. Further studies should investigate in depth the naturally occurring mechanisms that damage the tubules as well as those underlying the incidence of sperm granulomas.
Go to :
 XML Download
XML Download