

 PDF
PDF ePub
ePub Citation
Citation Print
Print


A well-adapted microbial pathogen, Salmonella enterica is a gram-negative, facultative anaerobic bacterium, which belongs to family Enterobacteriaceae. It was found to cause a variety of illnesses ranging from common food poisoning to severe typhoid fever [1]. Especially, salmonellosis is known as a main public health problem worldwide both in humans and domestic animals [23]. Human cases of salmonellosis were often associated with food [4], but frequently people got infected by handling infected animals and contaminated water and soil [5]. Children below the age of 5 years, elderly people and immunosuppressed patients were noted more susceptible to Salmonella infection than healthy individuals [6].
Quinolones are often used to treat invasive salmonellosis in humans and animals, but past studies encountered a significant number of clinical isolates resistant or with a reduced susceptibility to these antimicrobials [78]. Salmonella strains isolated from humans and animals were reported to have nalidixic acid resistance and considerably reduced susceptibility to fluoroquinolones [910]. Quinolone resistance among Salmonella spp. is generally associated with mutations in the genes encoding DNA gyrase (gyrA and gyrB) and topoisomerase (parC and parE) that is possessed in the quinolone resistance-determining region (QRDR) [11]. However, the production of Qnr proteins by plasmidmediated quinolone resistance (PMQR) genes was also documented as an another mechanism leading to quinolone-resistance [12]. These PMQR genes include; DNA gyrase protection from fluoroquinolone caused by a cluster of proteins encoded by qnrA, qnrB, qnrC, qnrD, and qnrS and a variant of aminoglycoside acetyltransferase capable of acetylating several fluoroquinolone compounds, aac(6')-Ib-cr [13].
Chelonians, consisting of turtles, tortoises, and terrapins, are kept in captivity for commercial purposes, and mostly as pets. So far, turtle keeping trend is continuously increasing but, the turtle-associated medical knowledge still keeps slow growing. However, pet turtles are known to harbor a variety of zoonotic bacteria, especially, in their digestive tract [1415]. Several previous studies have been conducted to assess the bacterial flora harbored by pet turtles and their environment [161718]. Turtle-borne Salmonella, in particular, has been well-recognized as a major cause of salmonellosis in humans [19]. Due to the concerns about the excretion of pathogenic Salmonella spp. the sale of small carapace turtles was banned in the United States' domestic pet market in 1975 by the Food and Drug Administration [20]. In the meantime, plasmid mediate high-level gentamicin resistance was detected in S. enteric isolates from pet turtles in Lousiana, USA [21]. Both antimicrobial-resistant and susceptible Salmonella spp. has also been isolated from Russian tortoises and spur-thighed tortoises [2223]. Another recent study recorded antibiotic-resistant S. enterica isolated from chelonians which were also harboring some virulence genes in their plasmids [24]. Besides, S. enterica could be isolated from several popular species of pet turtles and their environment indicating a potential risk of infection to turtle keepers [25].
Current study sought to characterize the S. enterica isolated from six commercially popular pet turtle species through the detection of quinolone susceptibility and screening the plasmid-mediated quinolone resistance genes (qnrA, qnrB, qnrS and aac(6')-Ib-cr) and chromosomal QRDR mutations in gyrA, gyrB and parC for a better understanding of a well-known turtle-borne pathogen.
Materials and Methods
Purchase of pet turtles
A total of thirty-five turtles comprising of 11 Chinese stripe-necked turtles (Ocadia sinensis), 8 yellow-bellied sliders (Trachemys scripta scripta), 6 river cooters (Pseudemys concinna concinna), 4 northern Chinese softshell turtles (Pelodiscus maackii), 3 western painted turtles (Chrysemys picta belli) and 3 common musk turtles (Sternotherus odoratus) were purchased from several pet shops and online markets in Korea. The purchased turtles had an average weight of 15±2 g, carapace diameter of 40±5 mm, and were under 4 weeks of age. Turtles were raised under laboratory conditions following the general husbandry method [26].
Isolation of Salmonella spp.
Fecal samples from turtles were enriched in tetrathionate broth (MBcell Ltd., Seoul, Korea) by incubation at 37℃ for 24 h. Enriched samples were streaked onto the selective culture media, brilliant green agar (BGA, MB cell Ltd., Seoul, Korea) and xylose lysine desoxycholate agar (XLD, MB cell Ltd., Seoul, Korea) and incubated at 37℃ for 24 hr. The isolates that showed the positive growth were presumptively identified as Salmonella spp.
Bacterial identification using 16S rRNA gene amplification and sequencing
Genomic DNA was extracted from presumptively identified Salmonella spp. isolates by Chelex-100 extraction method and PCR for 16S rRNA was performed using universal primers 12F and 1492R. Amplicons were sequenced and tested for the similarity using Basic Local Alignment Search Tool (BLAST) algorithm of NCBI database so as to confirm the species status.
Quinolone susceptibility testing
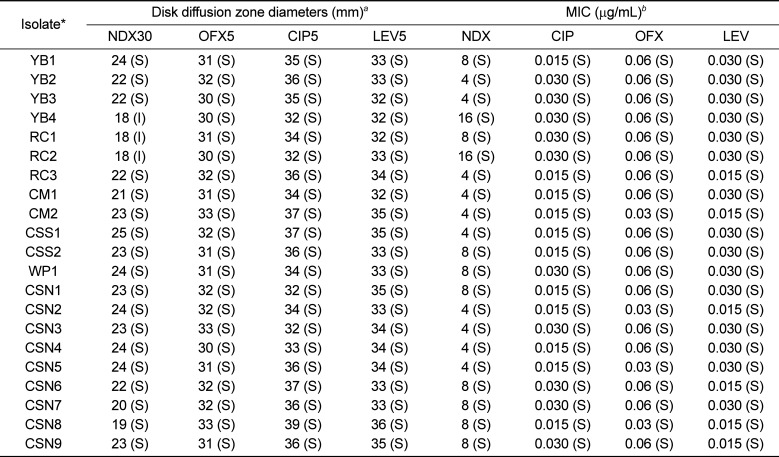
Twenty-one identified S. enterica isolates were selected for the study. Susceptibility pattern of the S. enterica isolates was investigated for nalidixic acid, ciprofloxacin, ofloxacin, and levofloxacin by disk diffusion test on Mueller Hinton agar (MBcell Ltd., Seoul, Korea). Minimum inhibitory concentrations (MIC) of nalidixic acid, ciprofloxacin and ofloxacin were detected by broth microdilution method. All susceptibility testing were conducted according to the recommendations of Performance Standards for Antimicrobial Susceptibility Testing; Clinical and Laboratory Standards Institute [27].
Detection of quinolone resistance genes
The presence of qnrA, qnrB, qnrS and aac(6')-Ib genes were detected using primers and PCR conditions listed in Table 1. Each PCR mixture was 20 µL total volume comprising of 10 µL of Quick Taq® HS DyeMix (TOYOBO, Japan) 1 µL of 10 pmol/µL each primer and 1 µL of the template. PCR products were checked in 1.5% agarose gel. Amplicons of aac(6')-Ib gene were purified using Expin™ PCR SV kit (GeneAll®, Korea) and were sent to Cosmogenetech Co. Ltd, Daejeon, Korea for sequencing. Acquired sequences were tested for the similarity in BLAST option in NCBI database in order to check the presence of aac(6')-Ib-cr gene variant.
Amplification of Quinolone Resistance Determining Region (QRDR) and detection of mutations
PCR for gyrA and parC regions were performed using primers and conditions described in Table 1. PCR was conducted in 50 µL volumes consisting of 20 µL of Quick Taq® HS DyeMix (TOYOBO, Japan), 2 µL of 10 pmol/µL each primer and 2 µL of the template. Amplicons were purified using Expin™ PCR SV kit (GeneAll®, Korea) and sent to Cosmogenetech Co. Ltd., Daejeon, Korea for sequencing. Acquired sequences were subjected to detection of mutations by comparison with published NCBI reference sequences. Analyzing and comparison of QRDR sequences were performed using Mutation Surveyor V5.0.1 (Softgenetics LLC, USA) software.
Phylogenetic comparison of gyrA sequences with published sequences
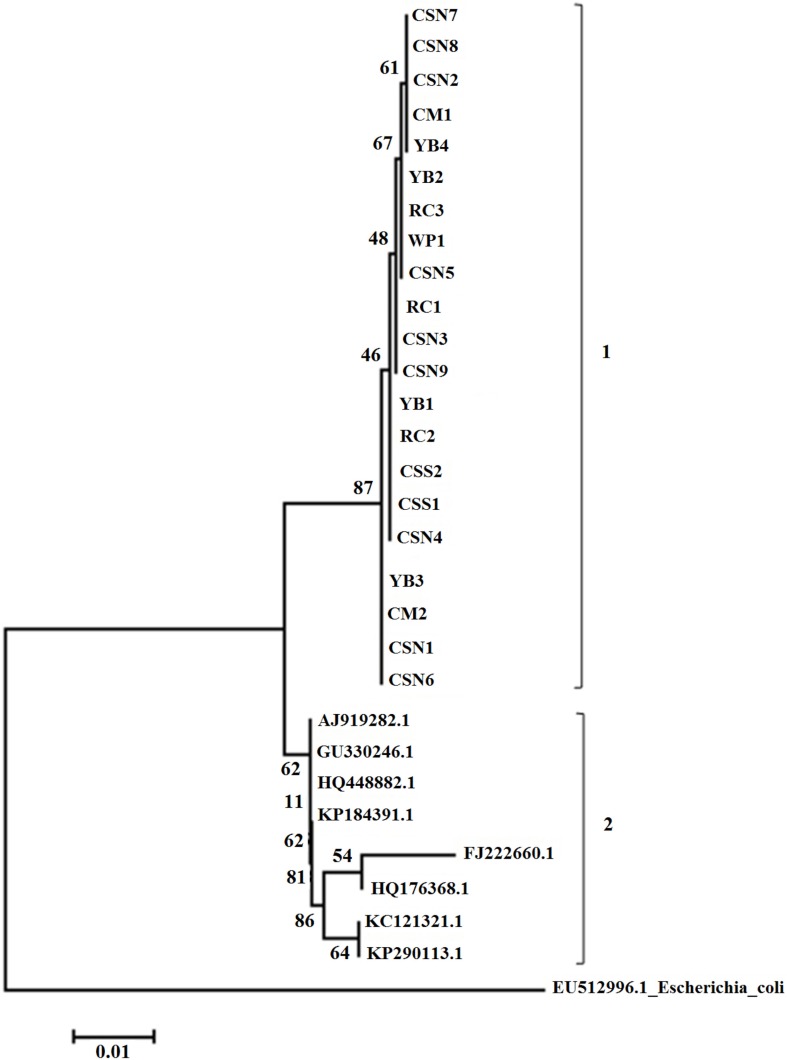
The sequences acquired for gyrA gene region were analyzed and a neighbor-joining phylogenetic tree was derived with 1000 bootstrap replications. For the analysis, eight previously published gyrA sequences of S. enterica subsp. enterica clinical isolates were obtained from the GenBank database (see Table 2 for the accession numbers and details) and MEGA6 sequence analyzing software was used for aligning and construction of the phylogenetic tree.
Results
Isolation and identification of Salmonella spp.
Twenty-one out of thirty-five isolates showed positive growth on BGA and XLD agar. A subsequent BLAST search after 16S rRNA sequencing indicated a 99-100% match to S. enterica sequences available in GenBank which confirmed their identity as S. enterica subsp. enterica.
Quinolone susceptibility testing
The results of the quinolone susceptibility testing are shown in Table 3. In disk diffusion test, only three isolates showed intermediate resistance to nalidixic acid and two of them showed reduced susceptibility in MIC. All tested strains are susceptible to other three fluoroquinolones; ciprofloxacin, ofloxacin, and levofloxacin both in disk diffusion test and MIC.
Detection of quinolone resistance genes (qnrA, qnrB, qnrS and aac(6')-Ib-cr) and QRDR mutations:
None of the isolates harbored qnr genes or aac(6')-Ib. Amplicon sequences of gyrA, gyrB, and parC did not show any specific point mutation in QRDR.
Phylogenetic comparison of gyrA sequences
Phylogenetic tree derived by analyzing and comparing the gyrA gene sequences obtained by current study and the published NCBI sequences is illustrated in Figure 1. Neighbor-joining phylogenetic tree indicated two distinct clads comprising first, current study isolates, second, quinolone-resistant isolates of human and animal origin.
Discussion
It is reasonable to deem that S. enterica is a pathogen which is able to cause serious infections in animals and human. In the meantime, it's intrinsic and acquired resistance to antimicrobials has also been well studied. Due to higher significance and implications counting current health concerns, studies associated with quinolone resistance, particularly genetics-based resistance, of S. enterica worth carrying out.
In this study, the majority of the isolates were susceptible to all tested quinolones except three isolates showing intermediate resistance to nalidixic acid. This outcome is in line with previous studies in which animal, especially reptile-borne S. enterica was examined. Nowakiewicz et al. reported S. enterica subsp. enterica strains isolated from Russian tortoises showing sensitivity to norfloxacin and other antibiotics [22]. S. enterica isolated from spur-thighed tortoises and tegu lizards exhibited 100% susceptibility to most of the tested antimicrobials including nalidixic acid and ciprofloxacin [2328]. In addition, Salmonella isolates from reptile feces were reported to show a higher rate of susceptibility to quinolones [29]. In contrary, clinical strains isolated from humans and poultry could resist nalidixic acid (88%) and were less susceptible to ciprofloxacin (23%) [30]. According to the personal communication with pet shops, the studied group of turtles was juvenile and aged less than 4 weeks. All the individuals were purchased soon after hatched, did not show any sign of disease and were not exposed to antibiotics. It could be the reason S. enterica subsp. enterica isolates to become more susceptible to quinolone antibiotics.
Previously, qnrA, qnrB, qnrS, and aac(6')-Ib-cr plasmid-mediated resistance genes were reported in quinolone-resistant, non-typhoidal Salmonella [36]. The Qnr protein induces resistance to nalidixic acid, thereby decreasing or limiting susceptibility to fluoroquinolones [31]. However, this study could not amplify any PMQR determinant while all the isolates were sensitive to tested quinolones. Studies reported S. enterica of poultry and human origin showing no aac(6')-Ib-cr in their plasmids despite they were resistant to quinolones [3033]. Moreover, Asai et al. [34] could detect qnrS among 0.08% of the S. enterica isolates of food-producing animals while Kim et al. [33] reported qnrB4 and qnrS1 in 3.2% of the isolates from poultry. Prevalence of qnr genes in S. enterica human clinical isolates was also reported as low in recent studies [3536].
In Salmonella, the most common point mutations associated with resistance to quinolones occur in the gyrA gene, resulting from substitutions of Ser-83 with Tyr, Phe, or Ala, and of Asp-87 with Asn, Gly, or Tyr [37]. In addition to above two sites, a recent study reported gyrA mutations of Ala-131 position in poultry and human isolates [30]. Another study could detect mutations in parC in addition to the gyrA mutations but in very low rate compared to gyrA [36]. Besides, Ser83-Phe substitution was detected in Salmonella spp. isolated from poultry [33]. Although some non-specific and silent mutations of the amplified region were noticed, current study could not identify any of above mentioned specific point mutations within the amino acid positions 67 to 106. Therefore, it is reasonable to assume that QRDRs of every isolate in this study is less mutated compared to highly resistant isolates.
Phylogenetic analysis of gyrA sequences produced a neighbor-joining tree showing 2 major clads referred to as 1 and 2. Interestingly, clad 1 comprised of isolates of the current study while clad 2 enclosing all clinical isolated both from human and animals. This outcome clearly interprets the genetic similarity of S. enterica subsp. enterica isolated from pet turtles and how they diverge from quinolone-resistant clinical isolates. Molecular phylogeny of Salmonella spp. has been investigated recently using gyrA region in which the antibioticresistant and sensitive isolates were separated into two different clusters [38].
In conclusion, it is plausible that the S. enterica subsp. enterica isolated from pet turtles are not resistant to tested quinolones and their QRDRs were genetically more conserved than that of quinolone-resistant strains. So it could be a good point for turtle keepers, but further studies concerned with more antimicrobials are highly recommended for a better understanding.


 XML Download
XML Download