

 PDF
PDF ePub
ePub Citation
Citation Print
Print


The immune system is the body's defense against foreign antigens, and produces many cells and molecules that distinguish and eliminate foreign matter. The immune mediators released from macrophages play significant roles in these processes [12]. After cells have been infected with bacterial endotoxin lipopolysaccharide (LPS), macrophages are activated, which generate numerous inflammatory mediators such as interleukin (IL)-6, tumor necrosis factor (TNF)-α, and nitric oxide (NO) [3]. IL-6, a soluble mediator, which is promptly produced in response to infection and tissue injury, contributes to host defense through the stimulation of acute phase response, hematopoiesis, and immune reactions by the regulation of cell growth, proliferation, and differentiation [4]. NO is a major component in the innate immune reaction of the host to various pathogens including bacteria, viruses, and fungi. It is involved in the pathogenesis and control of infectious diseases, tumors, and autoimmune processes [5]. B and T lymphocytes are major cellular components of the adaptive immune response and generate specific responses to maximally eliminate specific pathogens. Because low immunity or an immunocompromised state can alter the host susceptibility to pathogens, and this phenomenon was related to disease [6], it can be expected that raising immune ability would be very important in the maintenance of health.
The modulation and activation of the immune response to alleviate disease has been of interest where plants may represent an ideal adjunct to existing chemotherapy for the treatment of various diseases. Hence, many plants that can activate the immune system have been suggested [78910]. For example, the extract of Gelidium amansii, a red algae cultivated on the northeastern coast of Taiwan, activated macrophages through an increase in cell proliferation and enhanced the production of NO, TNF-α, IL-1β and IL-6 [2].
Mesembryanthemum crystallinum is a halophyte that changes from C3 photosynthesis to Crassulacean acid metabolism (CAM) in response to high photon flux density, drought, and salinity [11]. The plant is covered with trichomes called epidermal bladder cells (EBC), which is reflected by its common names: crystalline ice plant or ice plant [12].
Ice plant is capable of growing under high salt conditions through the synthesis of protective osmolytes and antioxidant molecules such as beta cyanins and other flavonoids [13]. Ice plant has been reported to improve the proliferation of keratinocytes and lipolysis [1415]. In addition, ice plant has been found to contain a significant amount of D-pinitol, which has been reported to show potent anti-oxidant [16], anti-diabetic [17], anticancer [18], and anti-inflammatory activity [19]. However, little is known about the immunomodulatory effects of ice plant. The present study was therefore designed to investigate the potential effects of IPE on the immune-enhancing function in vitro, in several immune cell lines, and in vivo, in mice administered IPE for 2 weeks.
Materials and Methods
Reagents
Ultrapure lipopolysaccharide (LPS) from Escherichia coli O111:B4 was purchased from InvivoGen (San Diego, CA).
Ice plant extract (IPE) preparation
A single seed of ice plant (Mesembryanthemum crystallinum) was planted in a plastic pot containing a sponge and germinated in a green house at 25℃ with an ordinary nutrient solution. One month after the seeds were germinated, plantlet samples were freeze-dried in a vacuum for a minimum of 72 h and ground into a fine powder using a mortar and pestle. One hundred grams of each powdered sample was extracted with 10 L of 70% ethanol for 4 h at 40℃ in a shaking incubator. After centrifugation to remove impurities, the ethanol of the supernatant was evaporated using a rotary vacuum evaporator and the concentrate was diluted with distilled water to a final volume of 1 L. After filtration through a 0.45-µm PVDF filter, the extract was ready for use in experiments.
Animal experiments
Seven-week-old male C57BL/6 mice were purchased from Koatech (Pyeongtaek, Gyeonggi-do, Korea). The animals were maintained for 1 week on a commercial pellet diet and then randomly divided into four groups (n=4), which were orally administered IPE (25, 125, and 250 µL/mL) or vehicle once a day for 2 weeks. All animal studies were approved by the Institutional Animal Care and Use Committee of Chonnam National University (Approval No. CNU IACUC-YB-2016-53).
Cell culture and treatment
Raji and Jurkat cells were purchased from American Type Culture Collection (Manassas, VA, USA). Raji and Jurkat cells are suspensions cell and the first continuous human cell lines from B lymphocytes and T lymphocytes, respectively. Both cell types were maintained in RPMI-1640 medium supplemented with 10% FBS and 1% penicillin/streptomycin in a 5% CO2 incubator at 37℃. Bone marrow-derived macrophages (BMDMs) derived from murine bone marrow were prepared as previously described [20]. Briefly, BMDMs were cultured in complete Iscove's modified Dulbecco's medium (IMDM, Gibco, Grand Island, NY, USA) supplemented with 30% L929 cell culture supernatant, 10% FBS, 1% sodium pyruvate, 1% MEM Non-Essential Amino Acids (MEM NEAA), and 1% penicillin/streptomycin in a 5% CO2 incubator at 37℃. After 3 days, 10 mL of fresh medium was added, and the cells were incubated for an additional 2 days.
Cell proliferation assay
Cell proliferation was measured by the EZ-Cytox (Daeillab service, Seoul, Korea) assay kit based on water-soluble tetrazolium salt (WST). Briefly, Raji and Jurkat cells were seeded in tripliate in a 48-well plate at a density of 8×105 cells/well and incubated for 24 h. Subsequently, the cells were treated with different concentrations of IPE. After 24 h, the WST reagent solution (10 µL) was added to each well. The plate was incubated for 3 h in a CO2 incubator and measured by using an ELx808 microplate spectrophotometer at a wavelength of 450 nm.
Measurement of cytokines
To determine the production of IL-6 and TNFα, BMDMs were seeded in triplicate in a 48-well plate at a concentration of 8×105 cells/well and treated with LPS (10 ng/mL) in the absence or presence of IPE (2.5, 5, and 10 µL/mL) for 24 h. The concentrations of IL-6 and TNF-α in culture supernatants were determined by using a commercial enzyme-linked immunosorbent assay (ELISA) kit (R&D Systems, Minneapolis, MN, USA).
Real-time quantitative polymerase chain reaction (qPCR)
iNOS gene expression levels were determined by real-time qPCR. BMDMs were treated with IPE (1, 2.5, 5 µL/mL) for 6 h, RNA was extracted using the easy-BLUE™ Total RNA Extraction Kit (iNtRON Biotechnology, Seongnam, Korea), and cDNA was prepared from 0.1 µg of RNA using ReverTra Ace® qPCR RT Master Mix (TOYOBO Bio-Technology, Osaka, Japan) in accordance with the manufacturer's instructions. Real-time PCR was performed using the SYBR Green PCR Kit (Qiagen GmbH, Hilden, Germany). The primer sequences for RT-PCR amplification were as follows: iNOS; sense, 5′-GCATTGGAAGTGAAGCGTTTC-3′ and antisense, 5′-GGCAGCCTGTGAGACCTTTG-3′, GAPDH; sense, 5′-CGACTTCAACAGCAACTCCCA CTCTTCC-3′ and antisense, 5′-TGGGTGGTCCAGGG TTTCTTACTCCTT-3′. PCR was performed in a Rotor-Gene Q real-time PCR system (Qiagen) using a two-step protocol of 40 cycles of 95℃ for 10 s followed by 58℃ for 45 s. The results were normalized using GAPDH as an internal control.
Nitric oxide assay
BMDMs were plated at a density of 8×105 cells/mL in a 48-well plate and incubated with or without INF-γ (100 ng/mL) in the absence or presence of various concentrations of ice plant extract for 24 h. Nitrite accumulation in supernatants was assessed using the Griess reaction [21]. An aliquot of the culture supernatant (100 µL) was mixed with equal volumes of Griess reagent [0.1% (w/v) N-(1-naphthyl)-ethylenediamine, with 1% (w/v) sulfanilamide in 5% (v/v) phosphoric acid] and incubated at room temperature for 10 min. The absorbance was measured at 577 nm by using a microplate reader, and a series of known sodium nitrite concentrations were used to construct a standard curve of absorption.
Isolation of cells from spleen
Spleens were removed aseptically from mice and placed individually into petri dishes containing 3 mL of complete RPMI 1640 medium. To prepare single cell suspensions, the spleens were chopped into small pieces and the spleen tissue was forced up and down through a 3-mL syringe, as previously described [22]. The suspension was transferred to a 15-mL conical tube containing 3 mL of complete RPMI 1640 and centrifuged at 1700 rpm for 10 min. The cells were re-suspended in 0.87% Tris-NH4Cl and incubated for 10 min at room temperature to lyse the erythrocytes. After two washes in complete RPMI 1640, the suspensions were adjusted to a final concentration of 5×104 cells/wells in complete RPMI 1640.
Flow cytometry assay
Flow cytometry, which simultaneously measures multiple characteristics of single cells at a rapid rate, was used in our experiment to detect accessory molecule expression on spleen cells after fluorescence activated cell sorting (FACS). In brief, splenocytes were harvested and washed with 0.5% BSA. The cells were stained with fluorescently labeled monoclonal antibodies (mAbs) to the mouse antigens of interest at optimal concentrations for an additional 30 min at 4℃. Antibodies (BD Biosciences, San Jose, CA) were directly labeled with the following fluorescent tags: fluorescein isothiocyanate (FITC) or phycoerythrin-Cyanine 7 (PE-Cy™7) for CD4 [GK1.5 and Rat (LEW) IgG2b, κ], CD8 [53-6.7 and Rat(LOU) IgG2a, κ], and CD19 [1D3 and Rat (LEW) IgG2a, κ]. In the experiments, CD4- PE-Cy™7 was used as a specific marker for T-helper (Th) lymphocytes and NK-T lymphocytes, CD8-FITC was used as a specific marker for cytolytic T lymphocytes and dendritic cells, and CD19-FITC was used as a specific marker for B cells. Appropriate isotype controls were always included. After centrifugation, the cells were fixed with 1% formalin, and 10000 viable cells per treatment (determined from light scattering profiles) were analyzed by using a BD FACSCalibur™ flow cytometer and CellQuest software (BD Biosciences).
Statistical analysis
The differences among the mean values for the different groups were evaluated, and all values were expressed as the mean±standard deviation (SD). Statistical calculations were performed using GraphPad Prism version 5.01 software (Graph-Pad Software). Cytokine production was analyzed by one-way ANOVA followed by the Bonferroni post-hoc test for multi-group comparisons. Differences were considered significant when P values were less than 0.05.
Results
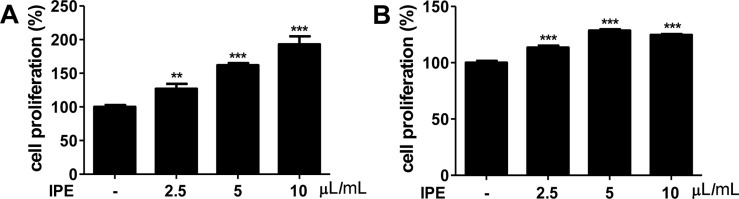
IPE enhances the proliferation of B and T cells in vitro
We first determined whether IPE promoted the proliferation of B and T cells. Raji (B cells) and Jurkat (T cells) cells were stimulated with various doses of IPE for 24 h and cell proliferation was determined by WST assay. IPE significantly promoted proliferation of Raji cells in a dose-dependent manner (Figure 1A). Moreover, the proliferation of Jurkat cells was significantly enhanced at all concentrations applied (Figure 1B).
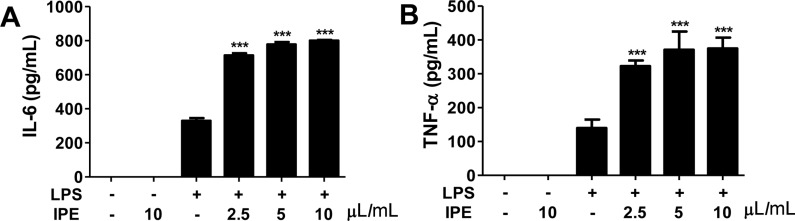
IPE enhances LPS-induced production of IL-6 and TNF-α in macrophage
We further sought to determine whether IPE induced cytokines production in macrophages. Bone marrow-derived macrophages (BMDMs) were stimulated with various doses of IPE in the range 0.1-50 µL/mL and levels of IL-6 and TNF-α in culture supernatants were determined by ELISA. IPE alone was unable to induce the proliferation of BMDMs and production of IL-6 and TNF-α (data not shown). However, IPE enhanced the LPS-induced production of IL-6 and TNF-α in BMDMs in a dose-dependent manner (Figure 2A, 2B). These results suggested that IPE may be able to cooperate with other immunostimulatory compounds to produce type I cytokines in macrophages.
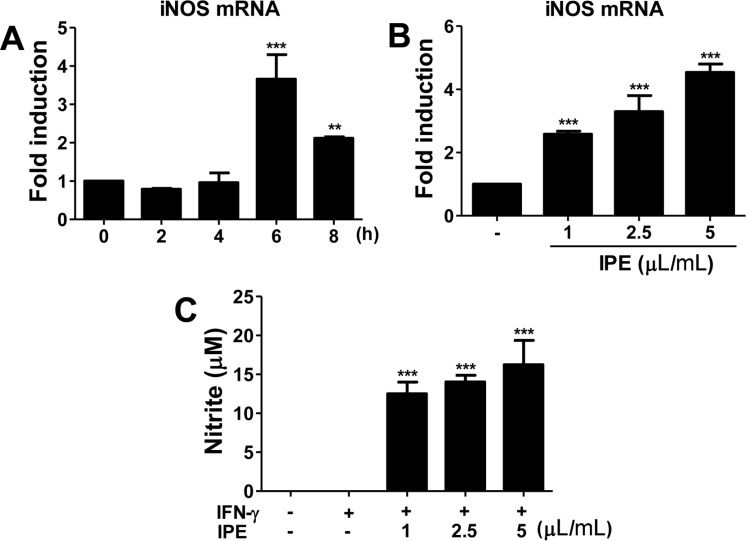
IPE leads to iNOS gene expression and NO production in macrophages
Nitric oxide (NO) is an antibacterial and antiviral effector of the innate immune system, and IFN-γ-induced inducible NO synthase (iNOS) is involved in the synthesis of NO from arginine. Therefore, we examined the effect of IPE on iNOS gene expression and NO production in BMDMs. IPE (5 µL/mL) induced optimal expression of iNOS mRNA at 6 h after treatment (Figure 3A). To determine dose-responsiveness, BMDMs were treated with the indicated doses of IPE for 6 h. iNOS mRNA expression was upregulated by IPE in a dose-dependent manner (Figure 3B). IPE-induced production of NO in the culture supernatant was determined by the Griess reaction. IPE alone did not produce a detectable level of NO in BMDMs (data not shown), although it did upregulate gene expression of iNOS. However, in the presence of IFN-γ, IPE induced NO production in BMDMs in a dose-dependent manner (Figure 3C).
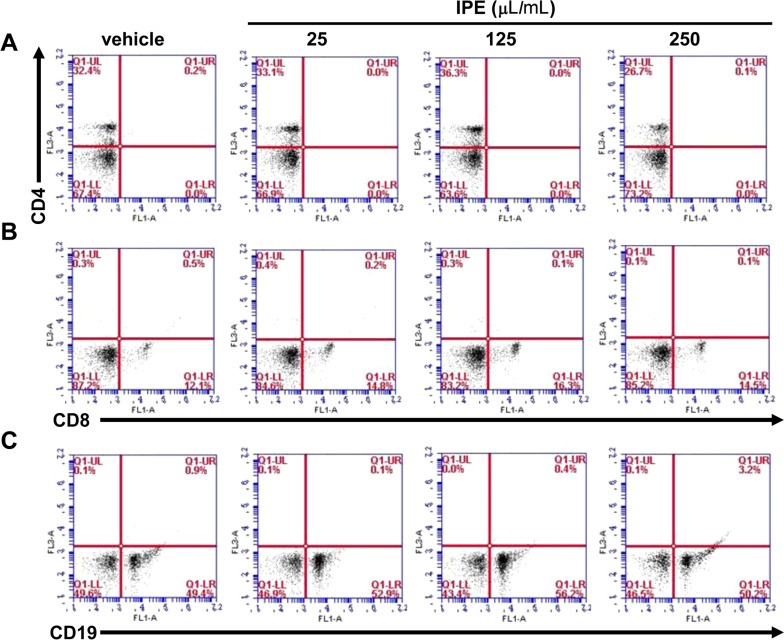
IPE enhances in vivo differentiation of splenocytes into CD4+, CD8+, and CD19+ cells in mice
To investigate the effect of IPE on in vivo immunomodulation, mice were orally administered IPE (200 mL) at doses of 25, 125, and 250 µL/mL for 2 weeks. The effect of IPE on splenocyte population in mice was measured by using flow cytometry to assess the expression of cell surface markers such as CD4+, CD8+, and CD19+. The percentage of CD4+ cells in the spleens was increased in response to IPE at doses of 25 and 125 µL/mL, but not at 250 µL/mL (Figure 4A). The population of CD8+ in the spleens was also increased in all groups administered with IPE compared with the vehicle group (Figure 4B). Differentiation into CD19+ cells was also increased in groups treated with IPE at 25 and 125 µL/mL, but was returned to original levels in the mice administered IPE at 250 µL/mL (Figure 4C).
Discussion
This study provided evidence of the in vivo immunomodulatory activities of IPE, which increased cell proliferation and the production of immune mediators. A variety of foods or plants have been reported to possess immune-enhancing activities through the activation of macrophages because the increased production of NO and cytokines such as IL-6 and TNF-α by the activated macrophages helps to clear pathogens [23]. IL-6 and TNF-α are pro-inflammatory cytokines and known to contribute to the inflammatory response and the development of various inflammatory diseases [2425]. TNF-α is essential in the containment of intracellular pathogens. It stimulates the recruitment of inflammatory cells to the area of infection and activates macrophages, which then phagocytose and/or kill mycobacteria and other pathogens [26]. In vitro studies have suggested several roles for TNF-α in the regulation of B and T cell proliferation and differentiation [2728]. IL-6 plays a protective role in the immune response against bacterial infection. IL-6 acts as a growth factor for mitogen-activated human B cells, stimulates their final maturation into antibody-producing plasma cells, and is involved in T cell activation and differentiation [2930]. In our study, IPE increased lymphocyte proliferation and enhanced the production of immune mediators, while the extract had no effect on macrophage proliferation. NO is a free radical synthesized from the amino acid L-arginine by the enzymatic activity of iNOS. The biological effects of NO have been identified as the regulation of vascular relaxation, platelet aggregation, and cellular respiration. In addition, it is known that NO can directly and indirectly modulate the immune response through the cytotoxic functions of macrophages against a variety of tumors and micro-organisms [31]. iNOS is absent from resting cells, and is expressed in response to pro-inflammatory cytokines such as IFN-γ, TNF-α, and LPS or microbial compounds [5]. Pyo et al. [32] reported that the immunomodulatory effects of Cimicifugae rhizoma extract comprise NO production and iNOS mRNA expression in macrophages. Therefore, the tumoricidal activity induced by C. rhizoma extract appeared to be mediated by the production of NO. Our study showed that IPE significantly upregulated IFN-γ-induced NO production and iNOS mRNA expression.
The cluster of differentiation (CD) is a protocol used for the identification and investigation of the cell surface molecules present on leukocytes. The two most commonly used CD molecules are CD4 and CD8, which are markers for T helper cells and cytotoxic T cells, respectively. T lymphocytes play a critical role in the development of the acquired immune response [33]. CD19, a human protein encoded by the CD19 gene, is critically involved in the establishment of intrinsic B cell signaling thresholds for antigen receptor-dependent stimulation [34]. Many plants extracts have been reported to exert immunostimulatory activity on different immune cells. For example, Han et al. showed that water extracts from various mushrooms promoted the population of CD3, CD19, CD4, and CD8 positive cells in the spleen [35]. To examine the effect of IPE on the population of T and B lymphocytes in the spleen, flow cytometric analysis was performed. Our data indicated that IPE increased the percentage of CD4, CD8, and CD19 positive cells populations in the groups administered 25 and 125 µL/mL, to a lesser extent, in the group administered 250 µL/mL. Overall, our findings indicated that IPE enhanced immune activities in vitro and in vivo.
In conclusion, the current study demonstrated that IPE was capable of efficiently enhancing immune functions. Overall, we confirmed that IPE possessed immunomodulatory functions, in which IPE may initiate innate immunity by an increase in macrophage proliferation, cytokine secretion, iNOS expression, and lymphocyte proliferation. Moreover, we suggested that IPE application, especially at concentrations of 25 and 125 µL/mL, increased the numbers of T and B cells in mice spleen. Our results support the idea that IPE has potential as an adjunctive immune-enhancing or immunomodulatory agent.
 XML Download
XML Download