

 PDF
PDF ePub
ePub Citation
Citation Print
Print


The persistent occurrence of acute lung injury (ALI) is a major clinical problem that leads to increased morbidity and mortality. ALI is characterized by pulmonary edema and severe acute inflammatory response; in particular, neutrophil accumulation results in dyspnea and death [1]. In the pathogenesis of ALI, excess neutrophil accumulation is facilitated by the complex interaction of inflammatory mediators [2]. Accumulated neutrophils can cause endothelial damage, enhance the permeability of the alveolar-capillary barrier, and release proinflammatory mediators, such as TNF-α and IL-6. Because these mediators exacerbate inflammatory responses, they are regarded as crucial markers of the acute phase of ALI [3456].
Lipopolysaccharide (LPS)-induced ALI animal models have been used to explore the mechanism of the pathogenesis of ALI and to investigate the therapeutic effects of the test material on ALI [7]. LPS, a major component of the cells wall of gram-negative bacteria, can induce pro-inflammatory cytokines and inflammatory cells [89]. LPS immediately stimulates the Toll-like receptor 4 (TLR4)-linked NF-κB pathway [10], which is related to not only a variety of physiological processes, but also to the development and progression of many inflammatory diseases [11]. NF-κB is activated by various stimuli and leads to the upregulation of many inflammatory mediators, such as TNF-α and IL-6, and inducible nitric oxide synthase (iNOS), which eventually induces pathophysiological responses that result in tissue damage [612]. Previous studies have shown that a reduction in the inflammatory responses, with a decrease in inflammatory cytokine and iNOS expression, was closely associated with the NF-κB pathway in in vivo and in vitro [1213].
The mugwort Artemisia argyi is used as tea and food in Korea [14]. A. argyi is a traditional medicine for the treatment of patients with abdominal pain, dysmenorrhea, and inflammation [15]. Recently, it was reported that A. argyi reduced allergic and inflammatory responses and oxidative stress [1617]. These properties were considered to be related to the amounts of phenolic compounds in A. argyi [1417]. We also recently demonstrated that A. argyi alleviates lung inflammation in an allergic asthma model and that it is the activity of Dehydromatricarin A in A. argyi [18]. However, there have been no studies on the effects of A. argyi against lung inflammation in an LPS-induced ALI model.
Therefore, we investigated the therapeutic effects of A. argyi and its active compound, dehydromatricarin A, on airway inflammation in an LPS-induced ALI model. In this study, we evaluated the production and expression of inflammatory mediators in LPS-induced lung tissue to clarify the possible mechanisms of action of A. argyi and dehydromatricarin A.
Materials and Methods
Experimental animal model
The experimental animal model used was designed by Shin et al. [13]. Male C57BL/6 mice (20-25 g) were purchased from Samtako Co. (Osan, Korea) and used when 8 weeks old. The mice were divided into seven groups: NC group, non-induced (control); LPS group, LPS-induced ALI; ROF group, roflumilast (10 mg/kg, P.O.)+LPS-induced ALI; AA-50 and AA-100 groups, A. argyi (50 and 100 mg/kg, P.O. respectively)+LPS; DA-10 and DA-20 groups; dehydromatricarin A (10 and 20 mg/kg, P.O. respectively)+LPS-induced ALI. LPS was administered by intranasal instillation of 10 µg in 50 µL PBS on day 5. Roflumilast, AA and, DA were orally administered for 7 days. Roflumilast is a PDE4 inhibitor used to treatment COPD. PDE4 inhibitor are known to possess anti-inflammatory effect. Seehase et al. suggested that roflumilast reduced pro-inflammatory cytokines such as TNF-α in LPS-induced acute lung inflammation [19]. Therefore, we used roflumilast as a positive control. The experiment was designed in accordance with the Institutional Animal Care and Use Committee of the Chonnam National University.
Measurement of inflammatory cells and cytokines in bronchoalveolar lavage fluid (BALF)
BALF collection was performed as previously described [15]. The animals received a tracheostomy under anesthesia and an endotracheal tube was inserted into trachea. BALF was obtained by filling the lung two times with ice-cold PBS (total volume: 1.4 mL). To count the inflammatory cells, 100 µL of BALF was centrifuged onto slides and stained by Diff-Quik® staining reagent (IMEB Inc., Deerfield, IL). The obtained supernatant was tested for inflammatory cytokines, including TNF-α, IL-6, and IL-β, by using commercial ELISA kits (R&D System, CA, USA) in accordance with the manufacturer's instructions. A quantitative analysis of cytokines was determined by the measurement of the absorbance at 450 nm using a microplate reader (Bio-Rad Laboratories, Hercules, CA, USA).
Measurement of inflammatory protein expression
To measure inflammatory protein expression, the lung tissue was homogenized with tissue lysis buffer supplemented with protease inhibitor (Sigma-Aldrich, Carlsbad, CA, USA) and the protein concentration was determined by using Bradford reagent (Bio-Rad Laboratories). Immunoblotting was performed as previously described [15] with the following primary antibodies: anti-iNOS (Santa Cruz, Santa Cruz, CA, USA), antip-NF-êB (1:1000 dilution; Abcam, Cambridge, MA, USA), anti-NF-κB (Abcam), anti-β-actin (Cell Signaling, Danvers, MA, USA). The band intensities of inflammatory proteins were measured by Chemi-doc (Bio-Rad Laboratories) and the relative expression values were computed by using IMT i-Solution software (IMT i-solution Inc., Vancouver, BC, Canada).
Histological analysis
The lung tissue was fixed in 10% (v/v) neutral buffered formalin. The tissues were embedded in paraffin, sliced into 4-µm sections, and stained with hematoxylin and eosin (H&E) solution to estimate inflammatory cell recruitment. Each slide was examined manually in a completely blinded manner using a light microscope (Leica, Wetzlar, Germany) equipped with 10× and 20× objective lenses and a 100× oil immersion lens. Images of 10 randomly selected non-overlapping areas per slide were captured with a digital camera (IMT i-Solution Inc.).
Results
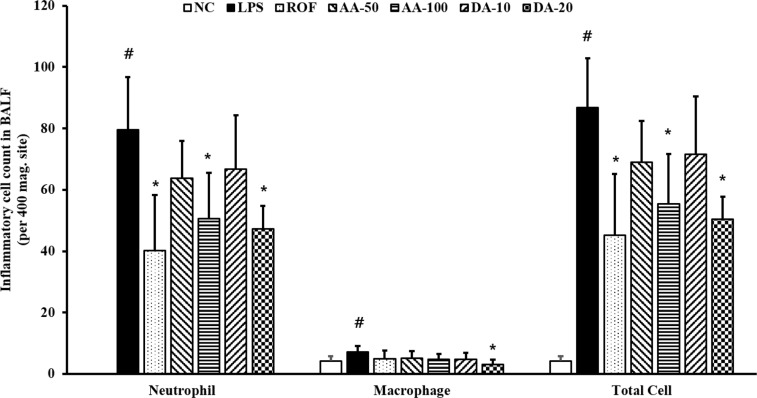
AA and DA decreased inflammatory cells in BALF
ALI-induced mice (ALI mice) showed a significantly elevated inflammatory cell count in BALF in comparison with normal, induced control mice (normal mice) (Figure 1). Among inflammatory cells, the neutrophil count in ALI mice was more increased than that in the normal mice. However, the inflammatory cell counts were decreased in AA-treated mice compared to that in ALI mice; the decrease was especially prominent at 100 mg/kg AA. Similarly, DA-treatment also clearly decreased the inflammatory cell count in comparison with that in the normal mice. AA (100 mg/kg) and DA (20 mg/kg) treatment resulted in a similar reduction in the inflammatory cell counts in BALF.
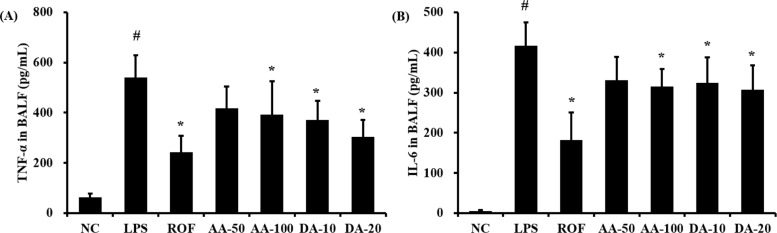
AA and DA reduced inflammatory cytokines in BALF
ALI mice significantly increased the level of TNF-α in BALF in comparison with that in normal mice (Figure 2A). AA treatment at 100 mg/kg significantly reduced the TNF-α level in BALF compared to that in the ALI mice; although a decrease was observed with 50 mg/kg AA, the difference was not statistically significant. The level of TNF-α was noticeably decreased in DA-treated mice compared to that in ALI mice. Similarly, the level of IL-6 in BALF was increased more in ALI mice than in the control mice (Figure 2B); however, AA and DA-treated mice exhibited markedly decreased IL-6 levels in comparison with the ALI mice, except for the 50 mg/kg DA treatment.
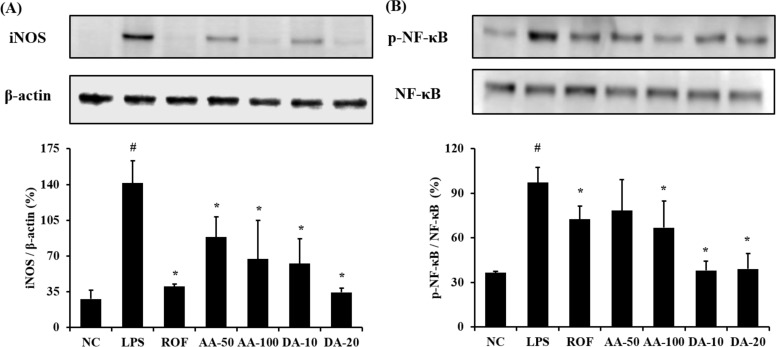
AA and DA decreased iNOS expression and NF-κB phosphorylation
ALI mice showed a marked elevation of iNOS expression in lung tissue compared with normal mice (Figure 3A). However, AA-treated mice significantly decreased the iNOS expression in lung tissue compared with ALI mice. In addition, DA-treated mice showed a notable reduction in iNOS expression compared with ALI mice. With regard to NF-κB phosphorylation, ALI mice displayed a notable elevation in lung tissue in comparison with normal mice, whereas AA- and DA-treated mice showed a significant reduction compared with ALI mice (Figure 3B). Although 50 mg/kg AA-treatment decreased the NF-κB phosphorylation compared with the ALI mice, the difference was not significant.
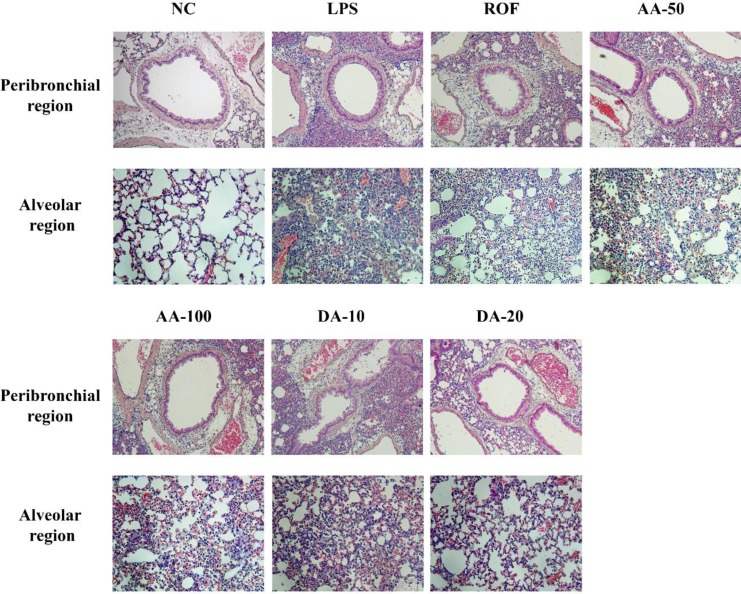
AA and DA attenuated inflammatory cell infiltration in lung tissue
In the histological examination of the lung tissue, ALI mice showed extensive inflammatory cell infiltration into peribronchial and alveolar lesions in comparison with normal mice. However, AA-treated mice attenuated the histological alteration induced by LPS instillation. Additionally, DA-treated mice clearly reduced the inflammatory cell infiltration into lung tissue in comparison with ALI mice.
Discussion
Acute lung injury (ALI) is a complex syndrome of intense pulmonary inflammatory response and the primary cause of death of intensive care patients [20]. In this study, we have explored the therapeutic effect of A. argyi (AA) and its active compound, dehydromatricarin A (DA), in an LPS-induced ALI mice model. AA-treatment in mice effectively suppressed the LPS-induced increases in inflammatory cell count and cytokines in BALF. AA also inhibited iNOS expression and NF-κB phosphorylation in ALI mice with a reduction in the inflammatory cell infiltration into lung tissue. Consistent with the results of AA-treatment, DA-treated mice also showed a significant reduction in inflammatory cell counts, cytokines, and pulmonary inflammation, which was accompanied by a reduction in iNOS expression and NF-κB phosphorylation.
ALI is characterized by the inflammatory responses that result from airway dysfunction and tissue remodeling in the respiratory tract. Inflammatory responses are regarded as an important defense system against exogenous harmful stimuli, but excess inflammation induces a loss of normal function and structural alterations owing to accumulated inflammatory cells and produces proinflammatory mediators, such as cytokines and chemokines [21]. In particular, among inflammatory cells, neutrophils are an essential component of the primary defense system; however, neutrophil accumulation into damaged lesions produced various proinflammatory cytokines and reactive oxygen species, which develop and aggravate ALI [22]. In this study, AA and DA treatment resulted in a significant reduction of the neutrophil count in BALF induced by LPS intranasal instillation. Furthermore, AA and DA treatment effectively attenuated the extensive inflammatory cell accumulation into lung tissue in ALI mice. These results indicated that AA has the potential to suppress neutrophil accumulation induced by LPS intranasal instillation.
TNF-α and IL-6 play key roles in the process of inflammation and stimulate the production of iNOS [23]. iNOS is a nitric oxide synthase (NOS) isoform; it is closely associated with NO production, which induces inflammatory responses [24]. In the pathogenesis of ALI, iNOS is considered a critical factor. Several studies have demonstrated that iNOS overexpression is related to the development and exacerbation of ALI owing to elevated NO and proinflammatory cytokine production in vivo and humans [2526]. In agreement with previous studies, LPS-induced ALI mice exhibited markedly increased TNF-α and IL-6 levels in BALF compared with the normal mice. However, AA and DA treatment significantly reduced the TNF-α and IL-6 levels by a decrease in iNOS expression in lung tissue in comparison with that in ALI mice. These results indicated that the therapeutic effects of AA may be related to the suppression of iNOS expression.
NF-κB is considered to be a limiting step in the inhibition of ALI development [252728]. In the inflammatory responses, NF-κB is activated by LPS stimulation and translocated into the nucleus via phosphorylation. Translocated NF-κB induces the transcription of inflammatory mediators, such as numerous cytokines, chemokines, and iNOS [2930]. Several reports have demonstrated that LPS-induced ALI increased the production of TNF-α and IL-6 and the expression of iNOS was increased via NF-κB signaling [63132]. Therefore, the suppression of NF-κB phosphorylation is considered an important therapeutic goal for the inhibition of ALI development. In this study, LPS-induced ALI mice clearly elevated NF-κB phosphorylation in comparison with the normal controls. In contrast, AA- and DA-treated mice significantly decreased NF-κB phosphorylation in comparison with ALI mice. These results indicated that the therapeutic effect of AA on ALI may be closely associated with the suppression of NF-κB phosphorylation.
In conclusion, our results showed that AA and DA effectively suppressed inflammatory cell count, cytokine production, and inflammatory cell accumulation in pulmonary inflammation in the LPS-induced ALI mice model. In addition, the effects of AA may be linked to the inhibition of NF-κB phosphorylation. Therefore, we suggested that AA has therapeutic potential for the treatment of LPS-induced acute lung injury.
 XML Download
XML Download