

 PDF
PDF ePub
ePub Citation
Citation Print
Print


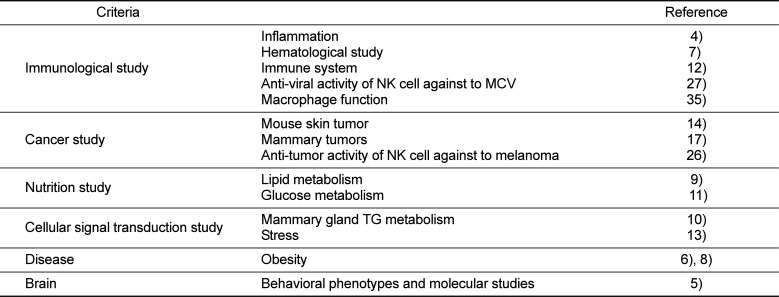
Experimental animal strains are useful animal resources. Such animal strains are generally involved in biomedical and behavioral experiments, and their use in experiments has contributed substantially to describing and elucidating biological mechanisms and metabolic pathways [1]. Specifically, inbred animal strains have been favored by researchers for improving the reliability and reproducibility of animal-based experiments. The origin of the inbred strain begins with the inbred mating experiment of guinea pig (Cavia porcellus) in 1906 by G. M. Rommel [2], and, since that time, various inbred strains of laboratory animals have been used. Among them, laboratory mice are preferred due to their short life span and ease of breeding, and a number of currently inbred mouse strains (e.g., C3H, C57BL/6 and CBA) have been developed. In addition, mouse genome sequencing has been completed, and the availability of such information has attracted much attention. Since the early 20th century when the first inbred mouse strain was established, a number of mouse types have been introduced [3]. Such mice are classified according to their respective mouse strains characteristics, such as hair color, blood parameters, biological behavior, immune response, response to stress, disease susceptibility (e.g., atherosclerosis, diabetes mellitus and its complications, and cancer), gene knockout, and a variety of trait-specific responses (Table 1) [4567891011121314].
Inbred mouse strains are produced after completion of 20 or more consecutive generations of sister-brother mating system [15]. Early inbred strains were developed for cancer and immunology studies, including some of the more famous early inbred hybrids; for example, C57BL/6, C57BL/10, C3H, CBA and BALB/c strains. Such inbreeding and the resultant variety of inbred strains have important roles in all areas of biomedical research by providing standardized animal models that enable independent researchers scattered around the world to obtain reproducible experimental results using the same material [16].
History of the C57BL/6 mice
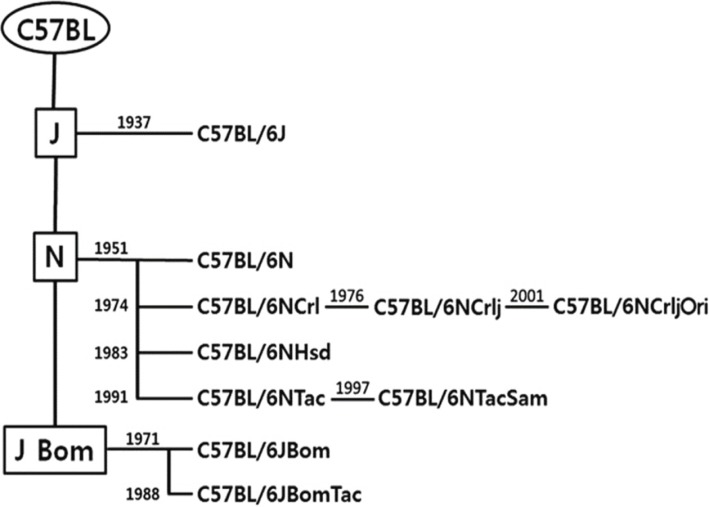
The C57BL mouse was established by C.C. Little in the 1920s and has been widely used as a universal stain and as a generic genetic background source of spontaneous or induced mutant mice. The C57BL/6 mouse strain is one of the more widely known inbred strains derived from the C57BL. Another widely known strain developed from C57BL is C57BL/10. The separation of C57BL/6 and C57BL/10 occurred in the 1930s. From the C57BL/6 strain, two of the more prominent substrains are C57BL/6J and C57BL/6N, which were developed by the Jackson Institute (JAX, Bar Harbor, Maine, USA) and the National Institutes of Health (NIH, Bethesda, MD, USA), respectively, in the 1940s and 1950s [117]. Substrains of the original inbred strain must meet three conditions in order to be recognized as a genetically different inbred substrain from the original inbred strain. First, the substrain is derived from 20th to 40th generation inbred mice. Second, generation branches are to be separated by more than 20 generations from the common ancestor. Third, there is to be a genetic difference between generation branches [18].
In the 1950s, C57BL/6J substrain mice were transferred from JAX to NIH and maintained by the NIH for several decades to produce a separate substrain known as C57BL/6N. Some C57BL/6N mice were then sent to commercial distributors Charles River Laboratories (Worcester, MA, USA) to establish C57BL/6NCrl, Harlan Laboratories (Latham Drive, Madison, WI, USA) to establish C57BL/6NHsd and Taconic Biosciences (Hudson, NY, USA) to establish C57BL/6NTac as well, some were sent to JAX to establish C57BL/6NJ (Figure 1). As required by the definition of a substrain, significant genetic variation may occur between these substrain. However, single nucleotide polymorphism (SNP) genotyping panels failed to detect a significant genetic difference among the Harlan, Taconic, or Charles River 6N substrains [1920]. Of course, current genetic testing has limitation, so a difference may not be detected. Therefore, it is necessary to use a substrain that is suitable for the research purpose and to use the same supply company in order to ensure the homogeneity of experimental animals.
C57BL/6 mice on immunological studies
Differences in the immunological responses of inbred mouse are of interest because they can be used to assess responses to pathogens and cell damage related to diseases and transplantation. C57BL/6 mice, in particular, are notable for their immunogenicity due to the initial strain development goal being their use in the study of cancer and immune responses. In general, the immune response is divided into an innate immunity and an adaptive immunity, and adaptive immunity is divided further into humoral and cell-mediated immunity. Innate immunity is broadly related to physiological barriers, such as the pH of the stomach, and anatomical barriers, such as the epithelial barrier and relatively immediate-acting phagocytes, natural killer cells, and the complement activation nonspecific defense system. Adaptive immunity is more specific than innate immunity and is mediated by B and T lymphocytes. However, the boundaries between innate immunity and adaptive immunity are not always distinguishable or mutually exclusive.
Cells associated with innate immunity include granulocytes neutrophils, eosinophils neutrophils, macrophages, and NK cells [21]. In general, these unique immune-related cells trigger nonspecific and acute immune responses, such as the onset of inflammatory reactions, and have been reported have major cellular roles. Moreover, their distribution within the immune system is important. Unlike myeloid lineage cells, NK cells have a lymphoid lineage and have an important role to function in antiviral and anti-tumoral responses and are particularly important as the primary donor of the interferon gamma cytokine [22].
Among the receptors that have important roles in the function of NK cells are killer cell lectin-like receptors (KLRAs) in humans. The Ly49 receptor in mice exhibits differences in specific receptor expression among inbred mice. For example, Ly49O and Ly49P are expressed in 129/J mice, whereas Ly49L is expressed in CBA/J and C3H/He mice. None of these are expressed in C57BL/6 mice. however, C57BL/6 mice do express receptors like Ly49H, and the differences in expression of these Ly49 receptors may contribute to differences in the activation of NK cells against tumors and viruses [232425].
As described above, C57BL/6 mice have been used since the early days of inbred strain establishment in studies related to response to antitumor and immunological response [26]. It is well known that the activity of NK cells, which have an important role in antitumor activity, is relatively high in C57BL/6 mice. C57BL/6 mice are also resistant to mouse cytomegalovirus through their expression of the LY49 receptor. These reports suggest that activation signals produced by NK cells may have an important role in the immune response differences between inbred strains. Results of activated receptor studies in rats are important when considering in the production of mutant mice for use in antitumor activity and immune studies [2728].
Adaptive immune responses are linked to native immunity, and lymphoid lineage B and T lymphocytes and their subtype cells are involved in adaptive immunity. For example, C57BL/6, unlike SJL/J, reverses the B:T cell ratio, suggesting that the ratio of B lymphocytes to T lymphocytes circulating in C57BL/6 mice is inversely proportional to other inbred strains such as SJL/J [29303132].
Cell-mediated T-lymphocytes consist of mediate T-helper cells, cytotoxic T cells, memory T cells, and other subtypes [3334]. Helper T cells are divided into TH1 and TH2 cells according to their mode of cytokine expression, which ultimately provides the immune response to the cell-mediated or to the humoral immune response by cytokines secrete and lead. C57BL/6 mice are characterized by the development of a high level of TH1 cells and a high TH1 response [35]. Another immune system, the humoral immune response, this is mainly driven by an antibody-mediated immune response guided by TH2, which in C57BL/6 is predominantly TH1-responsive and weaker than the cell-mediated responses.
Establishment of the C57BL/6NKorl stock
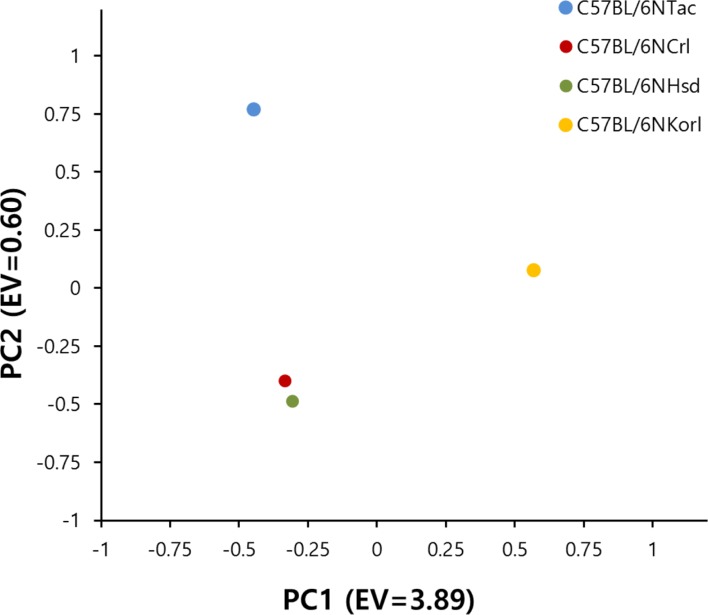
The results of research conducted by the Korean Food and Drug Administration for the localization of experimental animal resources produced a Korean local C57BL/6 stock, identified as C57BL/6NKorl, after inbreeding over 20 generations. C57BL/6NKorl was initially identified as C57BL/6J. However, that strain was confirmed as C57BL/6N after it was shown that an Nnt gene deletion did not occur, indicating no genetic differentiation from other cultivars. However, four single nucleotide polymorphisms (SNPs) proved C57BL/6NKorl to be a unique substrain. In 2015, the C57BL/6NKorl code was registered and certified by the Laboratory Animals Association (ILAR) under the National Institutes of Health, which manages the world's laboratory animal producer codes (Figure 2) [36].
Conclusions
Current inbred strains of mice are essential animal for laboratory research and such studies make clear contributions to many research fields. C57BL/6 mice is an inbred mouse strain originally developed for the study of antitumor activity and immunology, and is increasingly being used as an basic background strain. It is known that cell-mediated immunity and NK cell activity are of relatively high importance in determining immunological characteristics. In 2015, C57BL/6NKorl was established as a stock for N. The established C57BL/6NKorl is an independent stock showing genetic differences, based on SNP analyses, from other C57BL/6N. Studies into various characteristics of C57BL/6NKorl mice are expected to contribute to the use of this locally developed experimental animals and enhance the reliability and reproducibility of animal experiments.
 XML Download
XML Download