

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Histone acetylation and deacetylation is one of the important factors in the modulation of learning and memory, and of synaptic plasticity by histone deacetylases (HDACs, EC 3.5.1.98) [123]. HDAC proteins are grouped into four classes, of which class III consists of sirtuins (SIRTs), a family of NAD+-dependent proteins. Among SIRTs, SIRT2 is mainly found in the brain [456] and has been shown to be associated with the aging process and age-related neurodegeneration [4678]. Several lines of evidence indicate that SIRT2 inhibition facilitates neurite outgrowth in vitro and enhances neuroprotection in Parkinson's and Huntington's diseases [9101112]. However, Yuan et al. [13] recently found that SIRT2 inhibition accelerates neuronal damage in a model of traumatic brain injury. In addition, Chen et al. [14] found no effect of SIRT2 inhibition on brain injury such as amyotrophic lateral sclerosis and ischemic stroke. SIRT2 is closely related to AKT pathway in insulin as well as growth factor-responsive cells [15], and insulin receptor and growth factor receptors are highly expressed in the hippocampus [1617]. In contrast, HDAC inhibitors manipulate the epigenome and affects the expression of only 2% of mammalian genes [18]. In addition, HDAC inhibitors are essential agent for determining the role of histone acetylation in chromatin structure and function. Pharmacological inhibition by sodium butyrate, trichostatin A, or valproic acid show memory enhancement [1920] and increase the hippocampal neurogenesis [212223242526].
The hippocampus plays an important role in spatial memory and navigation. Newly generated cells in the subgranular zone of the dentate gyrus migrate into the granule cell layer and finally integrate as new neurons in the hippocampus in a lifelong process. Newly born neurons have a critical role in mediating pattern separation in memory formation and cognition in rodents [272829]. It is generally accepted that neurogenesis decreases with age [3031], but there are contradictory results with regard to SIRT2 expression in the brain. Braidy et al. [32] found that SIRT2 expression level was unrelated to age in the hippocampus of Wistar rats, while Kireev et al. [33] found a SIRT2 expression decrease in the dentate gyrus of old rats. In a previous study, we demonstrated that SIRT2 administration affects hippocampal functions such as memory formation, cell proliferation, and neuroblast differentiation [21].
In the present study, we investigated the effects of AK-7, a selective cell- and brain-permeable SIRT2 inhibitor [34] on novel object recognition memory, and on neurogenesis including cell proliferation and neuroblast differentiation in the dentate gyrus. We also looked at the relationship of AK-7 with sodium butyrate.
Materials and Methods
Experimental animals
Male C57BL/6 mice were purchased from Japan SLC Inc. (Shizuoka, Japan). They were housed in a conventional state under adequate temperature (23℃) and humidity (60%), controlled with a 12 h light/12 h dark cycle, and could freely access food and tap water. The handling and caring of the animals conformed to the guidelines established in order to comply with current international laws and policies (NIH Guide for the Care and Use of Laboratory Animals, NIH Publication No. 85-23, 1985, revised 1996), and were approved by the Institutional Animal Care and Use Committee (IACUC) of Seoul National University (Approval number: SNU-140404-5). All of the experiments were conducted with an effort to minimize the number of animals used and the suffering caused by the procedures used in the present study.
AK-7 treatment with or without sodium butyrate
AK-7 and sodium butyrate were purchased from Tocris Bioscience (Bristol, UK) and Sigma (St. Louis, MO, USA), respectively. The animals were divided into 3 groups (n=7 in each group): 1) vehicle-treated group (control), 2) AK-7-treated group without sodium butyrate (AK-7 group), and 3) AK-7-treated group with sodium butyrate (AK-7+SB group). Treatments were performed daily in 10-week-old mice for 3 weeks. AK-7 (20 mg/kg) was injected intraperitoneally and sodium butyrate (300 mg/kg) was administered subcutaneously because doublecortin (DCX) is exclusively expressed in immature neurons from 1 to 28 days of cell age [3536]. The concentration of AK-7 (20 mg/kg) was chosen because it was found to attenuate dopamine loss induced by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine in the striatum in the subacute paradigm [14]. In addition, the concentration of sodium butyrate (300 mg/kg) was selected as previous studies found that this dosage increases cell proliferation and neuroblast differentiation [212223].
Novel object recognition test
The test was performed in an acrylic box with three opaque walls and one transparent wall (45×45×30 cm). At day 20 of treatment, and 1 h after administration, the mice were placed in an open field and allowed to explore two identical objects for 5 min each and repeated once. Twenty-four hours after the open-field trial, one of the two familiar objects was replaced by a new one, and the mice were allowed to explore them for 5 min. In all experiments, relative exploration time was calculated as follows: relative exploration time in first and second day trials=100×[time observed each object (familiar or new)/time observing both objects]. The animals were sacrificed 2 h after the novel object recognition test.
Tissue processing
For histology, the animals from the control, AK-7, and AK-7+SB groups were anesthetized with 2mg/kg urethane (Sigma) and perfused transcardially with 0.1M of phosphate-buffered saline (PBS, pH 7.4) followed by 4% paraformaldehyde in 0.1M phosphate-buffer (pH 7.4). Brains were removed and post-fixed in a fixative for 12 h. Brain tissues were cryoprotected by infiltration with 30% sucrose overnight. Brain sections of 30-µm thickness were serially cut in the coronal plane using a cryostat (Leica, Wetzlar, Germany). Sections were collected in six-well plates containing PBS until further processing.
Immunohistochemistry
In order to obtain accurate data for immunohistochemistry, free-floating sections were carefully processed under the same conditions. For each animal, tissue sections were selected between 1.46 and 2.46 mm posterior to bregma by referring to the mouse atlas by Franklin and Paxinos [37]. Ten sections, 90 µm apart from each other, and were sequentially treated with 0.3% hydrogen peroxide in PBS and 10% normal horse serum in 0.05M PBS. They were then incubated overnight with diluted rabbit anti-Ki67 antibody (1:1,000; Abcam, Cambridge, UK) or goat anti-DCX antibody (1:50; Santa Cruz Biotechnology, Santa Cruz, CA, USA), and subsequently exposed to biotinylated horse anti-goat, or anti-rabbit IgG (diluted 1:200; Vector, Burlingame, CA, USA) and streptavidin peroxidase complex (diluted 1:200, Vector). Then, the sections were visualized by reaction with 3,3'-diaminobenzidine tetrahydrochloride (Sigma).
Ki67-, and DCX-positive cell counts were performed for each section of the dentate gyrus using an image analysis system equipped with a computer-based CCD camera (software: Optimas 6.5, CyberMetrics, Scottsdale, AZ, USA). Cell counts from all of the sections of all of the mice were averaged.
Statistical analysis
The data shown here represent the means±SEM. Differences among means were statistically analyzed by one-way ANOVA test, followed by Bonferroni post-hoc tests, in order to elucidate the effects of AK-7 with or without sodium butyrate on novel object recognition, cell proliferation, and neuroblast differentiation in mice. Statistical significance was considered at P<0.05.
Go to : 
Results
Effects of AK-7, with or without sodium butyrate, on object recognition
During the training period, control, AK-7, and AK-7+SB mice showed similar behavior, including similar spending of time exploring the two identical objects. During the test period, control mice spent more time exploring the new object than the familiar one, and the relative exploration time (67.52%) was significantly increased in this group compared to that in the control group spending time to identical objects (P<0.01). However, AK-7 mice spent less time exploring the new object compared to those in the control group, and the relative exploration time (57.01%) was not significantly different between familiar and new objects (P>0.05). AK-7+SB mice spent more time exploring the new object, and relative exploration time (63.06%) was also significantly larger than that for the AK-7 mice (P<0.05, Figure 1).
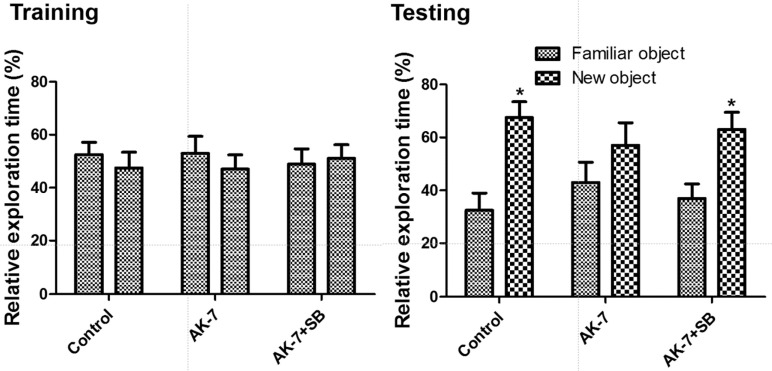 | Figure 1Effect of AK-7, with and without sodium butyrate, on exploration time of familiar vs. new objects in the novel object recognition test in mice (n=7 per group; *P<0.05, versus familiar object). Data for time of exploration for each object (same one, where one object was replaced by new one on the testing day) are presented as percentage of total exploration time. All data are shown as % exploration time±SEM.
|
Effects of AK-7, with or without sodium butyrate, on cell proliferation
In the control group, Ki67-positive nuclei were mainly detected in the subgranular zone of the dentate gyrus (Figure 2A) and the average number of Ki67-positive nuclei was 16.9 per section (Figure 2D). In the AK-7 group, a few Ki67-positive nuclei were observed in the dentate gyrus (Figure 2B), and the number of Ki67-positive nuclei was significantly decreased to 40.2% of the control group's (Figure 2D). In the AK-7+SB group, Ki67-positive nuclei were abundant in the dentate gyrus compared to the AK-7 group (Figure 2C), and the number of Ki67-positive nuclei was 71.0% of the control group's (Figure 2D).
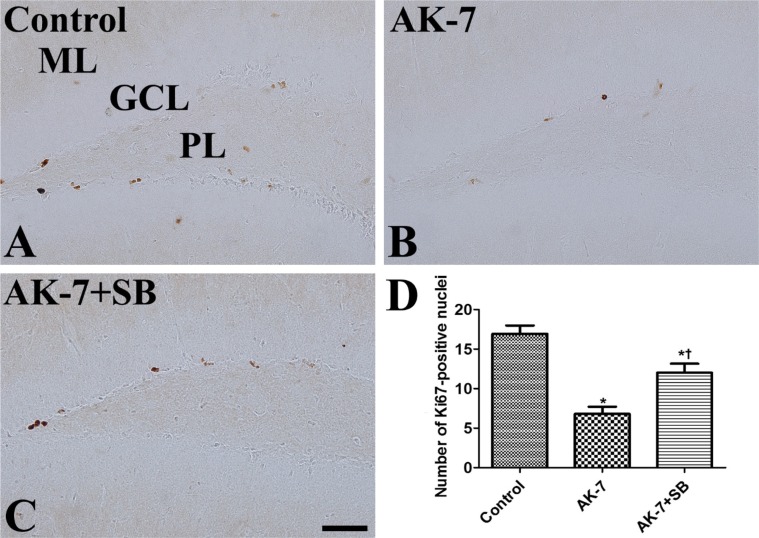 | Figure 2Immunohistochemistry for Ki67 in the dentate gyrus in vehicle-treated (control, A), AK-7-treated (B) and AK-7-treated with sodium butyrate (AK-7+SB, C) mouse groups. In the control group, Ki67-positive nuclei are detected in the subgranular zone of the dentate gyrus. Note that Ki67-positive nuclei are relatively few in the AK-7 group and are restored in the AK-7+SB group. GCL, granule cell layer; ML, molecular layer; PL, polymorphic layer. Scale bar=50 µm. (D) Number of Ki67-positive nuclei per section for each group (n=7 per group; *P<0.05, versus control group; †P<0.05, versus AK-7 group). Data are presented as mean±SEM.
|
Effects of AK-7, with or without sodium butyrate, on neuroblast differentiation
In the control group, DCX-immunoreactive neuroblasts were abundantly detected in the subgranular zone of the dentate gyrus (Figure 3A). In this group, DCX-immunoreactive neuroblast count was 131.4 per section (Figure 3D). In the AK-7 group, DCX-positive neuroblast count was significantly lower in the dentate gyrus to 47.0% of the control group's (Figure 3B, 3D). In the AK-7+SB group, DCX-immunoreactive neuroblasts were abundant in the dentate gyrus compared to the AK-7 group, and the number of DCX-immunoreactive neuroblasts was 81.3% of the control group's (Figure 3C, 3D).
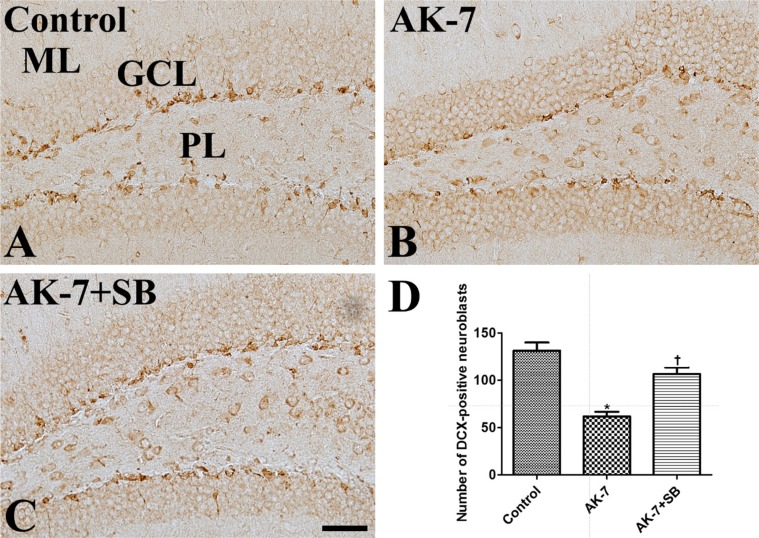 | Figure 3Immunohistochemistry for DCX in the dentate gyrus in vehicle-treated (control, A), AK-7-treated (B) and AK-7-treated with sodium butyrate (AK-7+SB, C) mouse groups. In the control group, DCX-immunoreactive neuroblasts are abundantly observed in the subgranular zone. Note that there are relatively few DCX-immunoreactive neuroblasts in the AK-7 group, and the reduction of DCX-immunoreactive neuroblasts is alleviated in the AK-7+SB group. GCL, granule cell layer; ML, molecular layer; PL, polymorphic layer. Scale bar=50 µm. (D) Number of DCX-immunoreactive neuroblasts per section for each group (n=7 per group; *P<0.05, versus control group; †P<0.05, versus AK-7 group). Data are presented as mean±SEM.
|
Go to :
Discussion
In a previous study [21], we demonstrated that the overexpression of SIRT by the PEP-1-SIRT2 fusion protein significantly causes memory deficits and reduces cell proliferation and neuroblast differentiation in the dentate gyrus. In addition, other researchers have shown that overexpression of SIRT2 inhibits neurite elongation [12] and SIRT2 knockdown enhances neurite outgrowth. In the present study, we investigated the effects of AK-7, a selective SIRT2 inhibitor permeable to the blood-brain barrier (BBB), on behavioral changes based on novel object recognition tests and neurogenesis based on cell proliferation and neuroblast differentiation. SIRT2 inhibition by AK-7 surprisingly also induced memory impairment and reduced neurogenesis as does SIRT2 overexpression. This result is supported by Liu et al. [38] who showed that SIRT2 inhibition by tenovin-D3 causes depression-like behavior and reduces the number of 5-bromo-2'-deoxyuridine (BrdU)-positive cells in the dentate gyrus of rats, while overexpression of SIRT2 by adenovirus significantly increases the BrdU-positive cells in the rat dentate gyrus [38]. Three-week long repeated treatment with MC1568, a histone deacetylase inhibitor, and 33i, a SIRT2 inhibitor, increased synaptic plasticity in the prefrontal cortex [39].
Results regarding SIRT2 function in the brain are contradictory. It has been reported that overexpression of SIRT2 decreases the survival of healthy neurons [40]. Conversely, SIRT2 inhibition prevents neuronal death from Parkinson's [1114] and Huntington's [41] diseases. However, SIRT2 inhibition has no effects on amyotrophic lateral sclerosis and ischemic stroke [14]. Knockdown of SIRT increases microglial activation and pro-inflammatory cytokines upon intra-cortical injection of lipopolysaccharide in mice [42]. In addition, inhibition of SIRT2 by AK-7 accelerates injury, BBB disruption, and microglial activation after controlled cortical impact [13]. A recent paper demonstrates molecular mechanisms of SIRT2 on the hippocampal neurogenesis, which is closely related to AKT pathway in insulin and growth factor-responsive cells [15]. Down-regulation of SIRT2 decreases AKT activation in insulin and growth factor-responsive cells, while overexpression of SIRT2 enhances the activation of AKT and its down-stream targets [15]. Collectively, these results suggest that the adequate levels of SIRT2 may be important to regulate the proliferation and differentiation of newly generated cells in the subgranular zone of the dentate gyrus.
The modulation of histone acetylation and deacetylation is critical for hippocampus-dependent object memory consolidation [24344]. In the present study, we investigated the effects of sodium butyrate on AK-7-induced reduction in novel object recognition memory and neurogenesis. In the previous studies, we and other groups demonstrated that sodium butyrate alters histone acetylation in the hippocampus [2345] and administration of sodium butyrate significantly increases the neurogenesis in the dentate gyrus of adult brain [212223242526]. In the present study, sodium butyrate increases the AK-7-induced reduction of novel object recognition memory, cell proliferation, and neuroblast differentiation. This result is supported by previous studies indicating that sodium butyrate ameliorates memory impairment caused by hippocampus-related diseases such as cerebral hypoperfusion, autism, and chronic mild stress [193946]. In addition, we demonstrated that sodium butyrate significantly increases cell proliferation and neuroblast differentiation in normal mice [23] and mice with brain ischemia [22]. The mechanisms involved were not elucidated, but it is possible that sodium butyrate reverses D-amphetamineinduced reduction in Krebs cycle enzyme activities including citrate synthase, succinate dehydrogenase, and malate dehydrogenase in the hippocampus [47]. Cranial irradiation was observed to alter Krebs cycle intermediates and glutamate/glutamine/GABA metabolism using in vitro
1H nuclear magnetic resonance spectroscopy [48]. It also significantly reduces neurogenesis [495051].
In conclusion, chronic inhibition of SIRT2 by AK-7 decreases novel object recognition memory and neurogenesis, and the supplementation of sodium butyrate increases the reduction these changes.
Go to :
 XML Download
XML Download