

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Inflammatory bowel disease (IBD), including ulcerative colitis (UC) and Crohn's disease (CD), are autoimmune diseases, characterized by chronic inflammation in the gastrointestinal (GI) tract, and recent reports indicated that the incidence and prevalence of IBD have increased worldwide [1]. Studies on pathogenesis of IBD have reported various factors that cause both types of IBD, including genetic susceptibility, environmental factors, infectious agents, and abnormality of immune system function [2]. IBD is accompanied by other diseases such as psychiatric disorders [34] and colon cancer development [56].
In the pathogenesis of IBD, the expression and secretion of pro-inflammatory cytokines, such as tumor necrosis factor-α (TNFα) and interleukin-1β (IL-1β), are increased and the expression of cytokines is regulated by nuclear factor-kappa B (NF-κB) [7]. In addition, dysfunction of the gastrointestinal barrier is a major step in the pathogenesis of IBD [8910]. The major components forming the gastrointestinal barrier are tight junction (TJ) proteins in the apical junction complex, which regulate paracellular permeability. It has been reported that the structure and function of TJ proteins are disrupted in IBD patients [11], which leads to intestinal bacterial translocalization into mesenteric lymph nodes [12].
There are several therapeutic strategies for remission of IBD, including use of anti-inflammatory medicines and immunomodulators, however, their clinical efficacy has been limited and accompanied by side effects, including fluid retention, insomnia and vomiting [13]. The overall remission rate of IBD patients with therapies is only approximately 50% [1415]. Among the therapeutic strategies, targeting the adoptive immune system, has been the most common for treatment of IBD, such as monoclonal antibodies against TNFα [1316]. However, treatment with the agents is successful only in less than one third of IBD patients and a proportion of patients experience loss of response to the agents [141718]. Therefore, development of new therapeutic strategies for treatment of IBD is needed.
Complementary and alternative medicines (CAMs) have recently been developed for treatment of IBD and, the use of herbal medicines is the most common for treatment of IBD [1920]. Herbal medicines have a number of beneficial effects, including anti-bacterial, anti-oxidant, and anti-inflammatory effects [19]. Pueraria L., a common Chinese herbal medicine, also has various beneficial effects, including anti-diarrhetic and antiemetic effects, as well as an anti-oxidant effect [2122]. Pueraria L. is rich in isoflavones, including peurarin, daidzein and genistein [21]. Isoflavones are known for their anti-inflammatory effect and anti-oxidant effect [23]. However, bioavailability of isoflavones is lower than that of their fermented forms, isoflavone aglycones [24].
In this study, for development of more effective herbal medicine for treatment of IBD, we generated the fermented Pueraria L. extract (fPue) and compared its anti-inflammatory effect with that of Pueraria L. extract (Pue). In addition, in vivo experiments were performed on a dextran sulfate sodium (DSS)-induced colitis model, which mimics human ulcerative colitis [25].
Materials and Methods
Generation of fPue
For generation of fPue, Pueraria L. was purchased from Omniherb Inc.; 400 g of that was crushed and boiled in 4 L of water at 121℃ for 30 minutes and the aqueous extract was filtered using Whatman filter paper 2 (GE Healthcare, Little Chalfont, United Kingdom). Dextrose (2%) and yeast extract (1%) were autoclaved and added to the Pue extract with 1% Lactobacillus brevis, aD110T, and the Pue extract was incubated at 30℃ for 5 days. Following incubation, the Pue extract was centrifuged at 12,000 rpm for 15 minutes, and the supernatant was filtered. The filtered extract was concentrated at 40℃ and 80 rpm, using a rotary evaporator (Eyela, Irvine, CA).
Animals
Specific pathogen-free female ICR mice (5 weeks old) were purchased from Orient Inc., Gyeonggi-do, Korea and the mice were acclimatized for 1 week prior to the experiment. All mice were maintained under 12 hours light/dark cycles at 22±5℃ and 60±5% humidity and provided with food (Orient Inc.) and water ad libitum. The information about components and calorie of the food is supported by Orient Inc. All experiments with animals were approved by the Institutional Animal Care and Use Committee at Lee Gil Ya Cancer and Diabetes Institute (IACUC No., LCDI-2014-0010).
Cell culture and treatment
Raw264.7 and Caco-2 cell lines were purchased from Korea Cell Line Bank, Seoul, Korea. The cell lines were cultured in 10% FBS-supplemented DMEM (WelGene, Daegu, Korea) with streptomycin (100 µg/mL)/penicillin (100 units/mL) at 37℃ in a humidified chamber with 5% CO2. Raw264.7 and Caco-2 cells were treated with Pue or fPue for western blot, RT-PCR and immunofluorescence analysis. For detection of NF-κB signaling by western blot, Raw264.7 cells were treated with Pue or fPue (0, 25, 100, 400 µg/mL) for 4 hours, followed by treatment with lipopolysccharide (LPS) for 1 hour. Total protein was isolated and western blot was performed. For determination of transcriptional level of cytokines in Raw264.7 cells, the cells were treated with Pue or fPue (0, 25, 100, 400 µg/mL) for 4 hours, followed by treatment with LPS for 3 hours. Total RNA was isolated using TriZol (Invitrogen, Waltham, MA) and RT-PCR was performed. Caco-2 cells were seeded in 12-well plates and cultured with or without a cover slip for immunofluorescence assay or western blot, respectively. The cells were incubated until they become 100% confluent, followed by incubation for an additional 7 days. The cells were treated with fPue (0, 25, 100, or 400 µg/mL) for 48 hours, followed by treatment with 2% DSS for 24 hours. Then, immunofluorescence assay and membrane fractionation for western blot were performed.
Induction of colitis by DSS
The mice were divided into 3 groups (7 mice in each group): (1) no treatment group; (2) 5% DSS-treated group; (3) 5% DSS with fPue (100 mg/kg)-treated group. DSS (MP Biomedical, Burlingame, CA) and fPue were dissolved in distilled water. Five percent DSS was administered to the mice in the drinking water for 7 days and normal distilled water was then supplied for 4 days for recovery (Figure 2A). During the recovery period, the mice were treated with fPue, or distilled water daily by gavage (Figure 2A). At the end of the recovery period, disease activity index (DAI) score was determined as described in the previous report [26] and necropsy was then performed.
Histological evaluation of colitis
The mice were sacrificed at the end of the recovery period. Following excision of the whole colon from anus to cecum, the length was measured. Approximately 0.5 cm of the distal colon was excised and frozen in liquid nitrogen for RT-PCR and histological analysis, and the remaining colon was Swiss-rolled and fixed in 10% formalin. The tissue samples were then paraffin-embedded and sectioned at 5 µm-thickness. The sections were stained with hematoxylin and eosin to evaluate the severity of colitis, according to three parameters described in the previous report [26]. The severity of colitis was scored blindly by 2 researchers.
Western blot
Twenty micrograms of each protein sample was used for western blot analysis. The primary antibodies are as followed: anti-rabbit ZO-1, anti-mouse Occludin, 1:1,000, anti-mouse Claudin-1 (Invitrogen, Waltham, MA); anti-rabbit p-P65 (Cell Signaling, Danvers, MA), anti-rabbit P65, anti-goat Lamin B (Santa Cruz, CA) and anti-mouse GAPDH (Merck Millipore, Billerica, MA).
Reverse transcription-PCR
Using 3 µg of RNA, cDNA was synthesized using the PrimeScript RT reagent Kit (TaKaRa, Shiga, Japan) as described in the manufacturer's instructions. RT-PCR was performed, using the primers listed on Supplementary Table 1 (Table S1).
Immunofluorescence analysis
The 5 µm-thick frozen colon tissue and cells on the slides were fixed in an acetone-methanol (1:1) mixture at -20℃. The slides were incubated with primary antibodies (anti-rabbit ZO-1, anti-mouse Occludin, anti-mouse Claudin-1; Invitrogen, Waltham, MA) overnight at 4℃ and secondary antibodies (anti-rabbit IgG FITC, Santa Cruz; Alexa Fluor 488 goat anti-mouse IgG; Invitrogen) for 2 hours at room temperature. After mounting the slides with VectaShield Mounting Medium with DAPI (Vector Laboratories, Burlingame, CA), the slides were sealed with nail polish and stored at -20℃ until observation, using a confocal microscope (Nikon Instruments Inc., Melville, NY).
Statistical analysis
Numerical data for all graphs were expressed as mean±standard deviation. Statistical changes in all data were determined using one-way analysis of variance (ANOVA). Scheffe was used for the post hoc test. The significance level was limited to 5% (P<0.05). SPSS 22.0 was used as a tool for statistical analysis.
Results
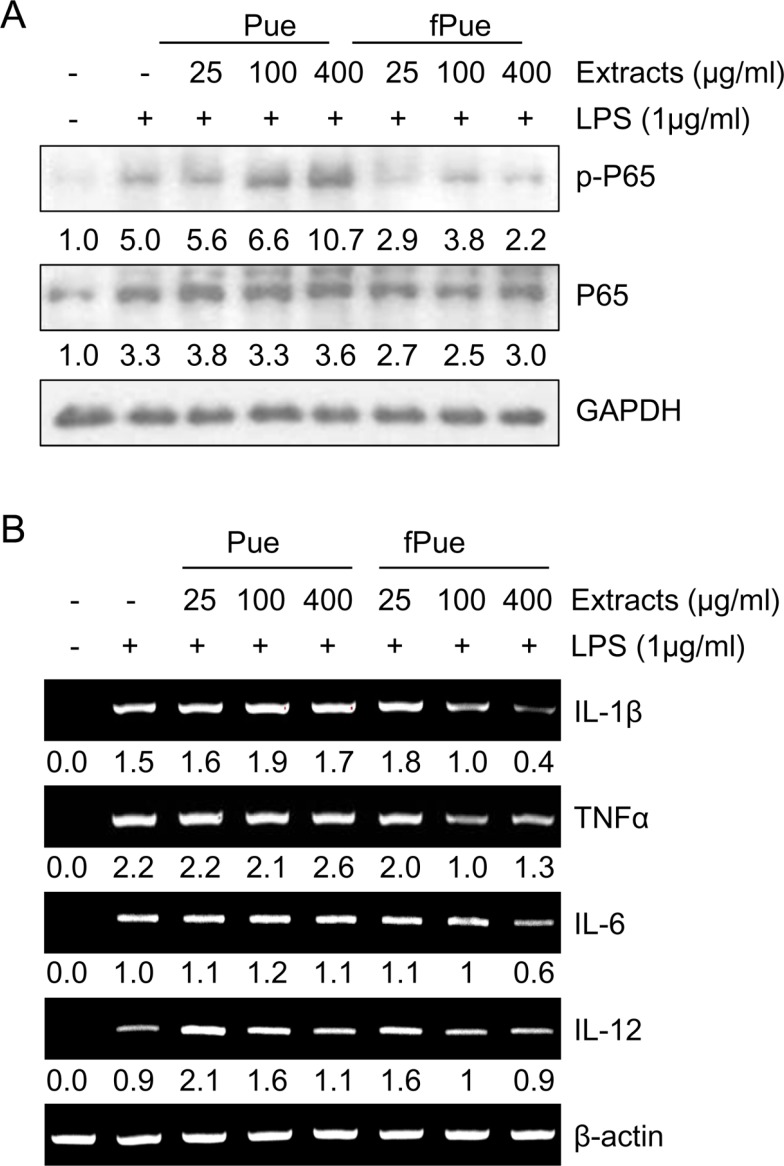
Anti-inflammatory effect of Pue is enhanced though fermentation in Raw264.7 cells
To determine whether fPue can enhance anti-inflammatory effect, compared to Pue, the activity of P65, a subunit of NF-κB, was examined in murine macrophage cell line Raw264.7. The cells were treated with Pue or fPue, followed by activation by LPS, which induces phosphorylation of P65 in Raw264.7 cells. Results of western blot showed that treatment with fPue significantly inhibited phosphorylation of P65 while Pue slightly inhibited that of P65 (Figure 1A). Activation of NF-κB leads to upregulated transcription of pro-inflammatory cytokines, including IL-1β, TNFα, IL-6, and IL-12. Treatment of LPS resulted in increased transcriptional levels of IL-1β, TNFα, IL-6 and IL-12 (Figure 1B). On the other hand, when pre-treated with fPue, the expression levels of the cytokines were decreased while treatment with Pue did not affect them (Figure 1B). These results suggest that fPue would improve the anti-inflammatory effect.
fPue has a therapeutic effect on DSS-induced colitis in vivo
Because fPue exerted an anti-inflammatory effect in Raw264.7 cell line (Figure 1), we examined whether fPue could improve DSS-induced colitis. DSS-induced colitis is characterized by disruption of epithelial structure and infiltration of inflammatory cells and loss of goblet cells [27]. To evaluate the effect of fPue on DSS-induced colitis, DAI was assessed by fecal analysis, to examine how much water and blood the fecal pellet contains and histological analysis was performed by H&E staining on the paraffin block sections to assess the severity of colitis. In the control group, DAI score was nearly 0 and that of the DSS-treated group showed a significant increase (Figure 2B), indicating that colitis was induced by DSS. On the other hand, DAI score of the DSS+fPue-treated group was significantly decreased, suggesting that fPue treatment improved the colonic function (Figure 2B). Histological analysis demonstrated that DSS treatment induced severe inflammation in mucosa, edema in submucosa and epithelial cell loss while they were significantly lower in the DSS+fPue-treated group, compared with the DSS+Vehicle-treated group (Figure 2C, 2D). In addition, alcian blue staining showed that while goblet cells were lost in the DSS-treated group, they were restored by fPue treatment (Figure 2E). These results suggest that treatment with fPue resulted in amelioration of colonic inflammation and promoted recovery of epithelial structure in the colonic mucosa, following recovery from diarrhea.
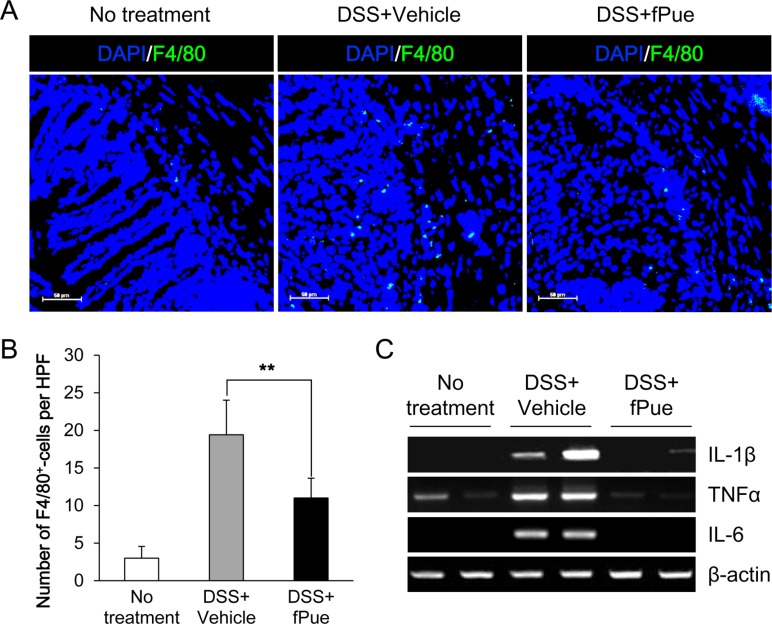
fPue treatment reduced infiltration of macrophages and pro-inflammatory cytokines in DSS-induced colitis
In a DSS-induced colitis model, inflammatory cells, mainly macrophages, are infiltrated into submucosa and lamina propria of the inflamed colon [1628]. To determine whether the number of infiltrated macrophages is decreased by treatment with fPue, immunostaining was performed for F4/80, a marker for macrophages, on frozen sections of colon tissue. Immunofluorescence analysis revealed that while the number of F4/80-positive cells present in mucosa and submucosa of the inflamed colon was significantly increased, that of F4/80-positive cells was markedly reduced in the colon of the DSS+fPue-treated group (Figure 3A, B), indicating that macrophage infiltration was reduced by fPue treatment. In addition, decrease in transcriptional expression of pro-inflammatory mediators, IL-1β, TNFα and IL-6 was observed by performing RT-PCR (Figure 3C).
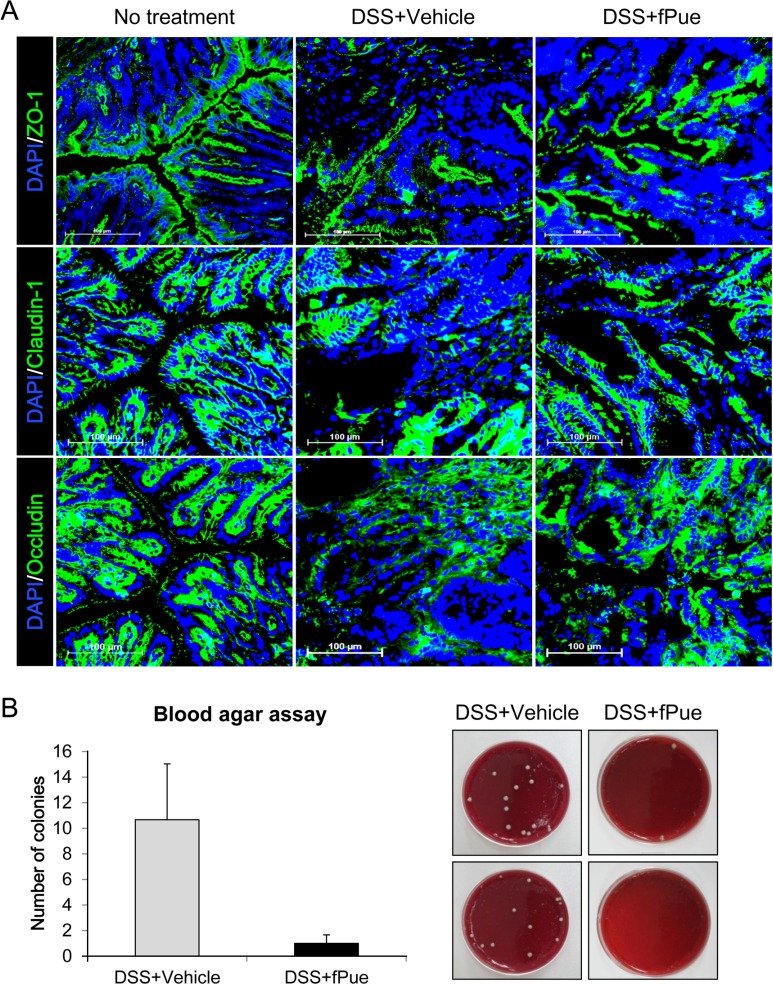
DSS-induced disruption of intestinal epithelial barrier was recovered by treatment with fPue
As previously mentioned, epithelial dysfunction is a major aspect in the pathogenesis of IBD. To determine whether the epithelial barrier, disrupted in DSS-induced colitis, is recovered by treatment with fPue, the expression levels and the arrangement of ZO-1, Claudin1 and Occludin, were examined by immunofluorescence assay, using cryosections of colon tissue. As shown in Figure 4A, all three TJ proteins were expressed along with the inner lining of the columnar epithelium of normal colon tissue. While the expression levels of TJ proteins were decreased and their architecture was disrupted in DSS-induced colitis, they were recovered as DSS-induced colitis was ameliorated by fPue treatment (Figure 4A). In addition, to investigate whether the physiological function of the epithelial barrier was recovered, bacterial translocalization was examined by blood agar assay, using cells from mesenteric lymph nodes. As shown in Figure 4B and C, bacterial translocalization was induced as the epithelial barrier was disrupted by DSS treatment. On the other hand, in the DSS+fPue-treated group a decreased number of bacteria was present in the lymph node (Figure 4B, C), indicating that bacterial translocalization was reduced as disrupted tight proteins were rescued by fPue treatment.
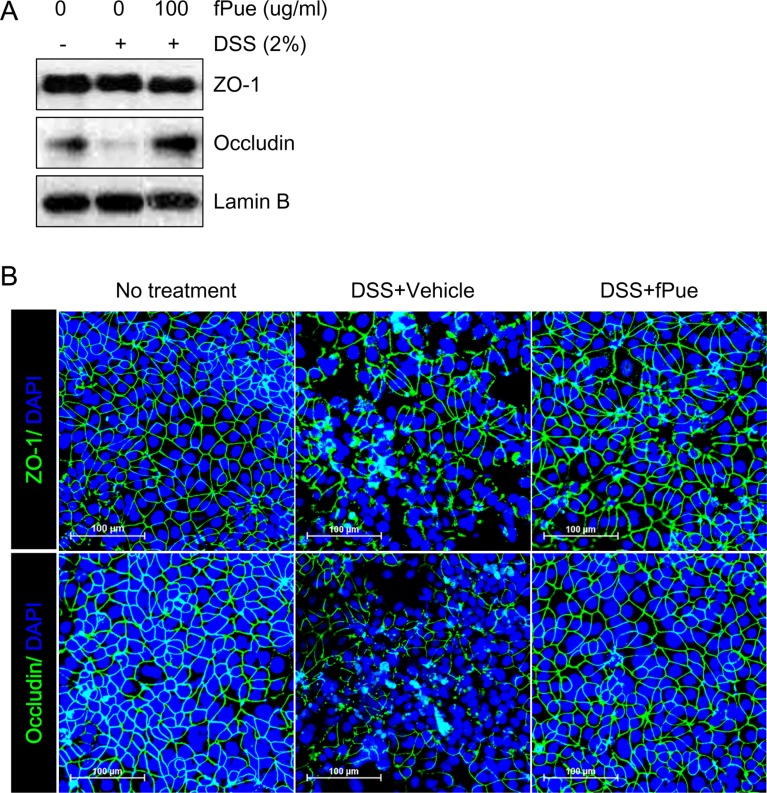
fPue directly recovers DSS-induced TJ damage in Caco-2 cells
To determine whether DSS-induced disruption of TJ would be recovered by fPue treatment, an in vitro experiment was performed using colonic epithelial cell line Caco-2. Caco-2 cells form a monolayer and TJ between the cells, and the TJ is disrupted by DSS treatment [29]. Consistent with the previous study, treatment of Caco-2 cells with DSS resulted in a decrease in protein expression of Occludin, which was detected by western blot, while that of ZO-1 did not change (Figure 5A). Although ZO-1 expression level was not affected by DSS, its architecture was disrupted as well as that of Occludin (Figure 5B). However, fPue treatment prevented disruption of the architecture of ZO-1 and Occludin (Figure 5B). These data suggest that fPue could have a protective effect directly on intestinal TJ.
Discussion
Previous studies have reported that traditional herbal medicines are the most commonly used CAM worldwide for treatment of IBD because of their efficacy and safety [1920]. However, because not all herbal medicines are safe and they are incomplete for treatment of IBD, improvement of the safety and efficacy is necessary. The fermentation process may be a useful tool for elimination of toxicity of herbal medicines and to improve their bioavailability [24]. Even though Pueraria L. is used traditionally in treatment of diarrhea, to the best of our knowledge, in vivo experiments, using fermented form of Pue on a DSS-induced acute colitis model have not been reported. Therefore, we aimed to investigate the differences in anti-inflammatory effect between Pue and fPue in vitro and to examine the therapeutic effect of fPue in vivo, using a DSS-induced acute colitis model in mice.
NF-κB signaling pathway is the major pathway that regulates expression of pro-inflammatory cytokines such as IL-1β, TNFα, IL-6 and IL-12, which are involved in development of IBD [16]. Therefore, inhibition of pro-inflammatory cytokines may lead to remission of IBD. In Figure 1, as Raw264.7 cells were treated with fPue, followed by LPS, phosphorylation of NF-κB and the expression of pro-inflammatory cytokines were reduced, compared to cells treated with LPS only. However, the treatment with Pue had a limited anti-inflammatory effect (Figure 1), suggesting that fermented form of Pue, rather than Pue itself, has an anti-inflammatory effect.
One of the major aspects in pathogenesis of IBD is infiltration of inflammatory cells, mainly macrophages [1628], which leads to production of pro-inflammatory cytokines. In our results, histopathological analysis showed that the number of infiltrated inflammatory cells was diminished in mucosa and submucosa of colon in the DSS+fPue-treated group, compared with the DSS+Vehicle-treated group (Figure 2, 3). Consistent with in vitro data, the transcription of pro-inflammatory cytokines was suppressed by fPue treatment (Figure 3C). This reduction of cytokines may have been resulted from a reduced number of macrophages infiltrated into colonic epithelium. In vitro data shown in Figure 1 also suggest that fPue could have a suppressive effect on production of pro-inflammatory cytokines in macrophages as it inhibits NF-κB activation. However, it remains unclear how fPue reduced macrophage infiltration and its cytokine production.
Another aspect of IBD is dysfunction of gastrointestinal barrier by destruction of TJ in colonic mucosa [8910]. The gastrointestinal barrier is responsible for uptake of nutrients, ions, and water, and for preventing translocation of pathogens across tissue [11]. Our in vivo experiment demonstrated that as acute colitis was induced by DSS administration in mice, the structure of the TJ complex was disrupted (Figure 4A). In contrast, the disrupted TJ complex was restored by fPue treatment (Figure 4A). In addition, while bacterial translocalization was induced due to disrupted TJ complex, bacteria were not translocalized into mesenteric lymph nodes as the TJ complex was recovered by treatment with fPue (Figure 4B, C), indicating that the function of intestinal barrier was recovered. Interestingly, treatment with fPue inhibited collapse of TJ proteins (Figure 5), suggesting that fPue may has a direct effect on recovery of the intestinal barrier function in acute colitis by rescuing TJ protein distribution.
In conclusion, in vitro experiment with Raw264.7 suggested that fPue has a more beneficial effect on inflammatory activity than the extract itself. The anti-inflammatory effect of fPue was also demonstrated in an in vivo experiment. fPue ameliorated DSS-induced acute colitis and recovered gastrointestinal barrier function. In addition, there was no side effect in the use of fPue, such as weight loss or liver injury (data not shown). However, investigation to determine whether there are additional beneficial effects of fPue is needed. In this study, we suggest that the fPue has a therapeutic potential for treatment of IBD without undesirable effects.

 XML Download
XML Download