

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Parkinson's disease (PD) is one of the most common neurodegenerative movement disorder, and affects ~2% of the world's population aged over 65 [1]. It is characterized by insufficient production of dopamine (DA) from the substantia nigra (SN) area of the brain, leading to asymmetric onset of bradykinesia, resting tremor, rigidity and postural instability. Pathologically, the hallmarks of PD are evolutional nigrostriatal DA neurodegeneration in the brain and the presence of cytosolic filamentous inclusions known as Lewy bodies (LBs) and Lewy neurites in surviving nigral DA cells [234]. Recent findings indicate that the mechanisms of DA neuron degeneration and death have been linked to mitochondrial dysfunction, oxidative stress, inflammation, and apoptosis [25]. In addition, PD also affects many other brain regions, such as the dorsal motor nucleus of the vagus, the nucleus basalis of Meynert, and the locus coeruleus [67].
PD is a result of complex interactions between environmental and genetic factors associated with a pathogenic mechanism and thus cannot be well studied using simple in vitro models [8]. For the past several decades, animal models of PD with their own strengths and weaknesses have been widely used to investigate the pathogenesis as well as possible innovative therapeutic approaches for this neurodegenerative disorder [9]. Experimental models of PD should reflect pathological, biochemical, and clinical features of PD including the lesions in both DA and non-DA systems. Although none of the current models show all the features of PD, animal models have contributed significantly to our current understanding of the pathological mechanisms of the disease and to the development of new therapeutic strategies in PD [10]. In this review, we examine the use of the common marmoset (Callithrix jacchus) (Figure 1A) treated with systemic or local administration of neurotoxins as an animal model to bridge the gap between rodent studies and clinical applications for the purpose of studying the pathogenesis and evaluating novel therapies for PD.
Characteristics of the Marmoset
The marmoset is a small New World primate that is originally from the Amazon basin of Brazil [11]. It is a relatively small animal, with an average height of 20-30 cm and weigh of 200-600 g (10-15-fold less than the 5-10 kg macaque). According to the United States National Research Council's "Guide for the Care and Use of Laboratory Animals", a small cage (Figure 1B) with a minimum height of 76.2 cm and a minimum floor area of 0.20 m2 is recommended for a breeding pair of marmosets. According to the European guidelines, marmosets should be kept in controlled facilities (23-28℃, 45-70% humidity, and 12 h light/dark cycle) since the animals originate from tropical rain forests [12].
The marmosets eat fresh fruit, bread, eggs, and nuts, and have high protein intake. Along with biscuits and condensed milk as remuneration food, a commercial diet must be supplemented with vitamin D3 since they require a large amount of vitamin D3 (e.g. "New World Primate Diet" by Harlan Teklad) [12]. The marmosets have high reproductive efficiency, with similarities to the human ovarian cycle (approximately 28 days), an early onset of puberty (around 1.5 years old), a relatively short gestation period (145-148 days), a relatively large litter sizes (2-3 offspring per delivery), and a relatively large number of deliveries (twice a year) [11]. They live in stable families of 1-2 breeding females, a breeding male, their offspring, and their adult relatives [13]. For toxicokinetic or clinical pathology studies, blood samples are usually collected via the femoral vein and occasionally the tail vein. Less than 15% of the circulating blood volume is recommended for blood collection within one month for the marmoset [12].
Because of its small size, ease of handling, and unique biological characteristics [14], the marmoset has become an important primate model in various areas of biomedical research such as neuroscience, toxicology, reproductive biology and regenerative medicine [15]. Importantly, the use of marmosets can lead to significant reductions in material requirements due to its small size [12].
Marmoset Model of PD
Transient parkinsonian-like states have been generated in various animal species from drosophila [16], to mice [17], rats [18], cats [19], minipigs [20], sheep [21], New World [222324], and Old World monkeys [4]. For many researchers, the mouse is a popular choice for behavioral assessments and screening for the effects of drug treatments due to a lack of resources and trained personnel for the monkey model. Monkeys have many similarities to humans in terms of developmental processes, brain anatomy/function, and social behaviors, hence, research on monkeys play an important role in the preclinical development process between rodent studies and controlled clinical trials [252627]. In particular, the use of the marmoset monkey requires less ethical justification than the larger "Old World monkeys" [12]. For this reason, there has been increasing interest in the marmoset monkey as a popular monkey species for the development of novel treatments for PD such as neurotrophic factors [28], DA reuptake inhibitors [29], and neurotransplantation [30].
Current animal models of PD include genetic and neurotoxic models. The genetic models are created primarily based on genes identified in potential mechanisms involved in the onset and propagation of PD in humans [931]. Over-expression of proteins such as á-synuclein and DJ-1 using viral vectors results in great practical significance of PD symptoms, leading to preclinical evaluations of various therapies for PD [932]. Recently, several genetically modified non-human primate (NHP) models were developed through the introduction of exogenous genes into NHP genomes or the alteration of endogenous NHP genes [3334]. This progress in knowledge and technology enable the production of transgenic marmoset models with clear PD phenotypes, which will have great practical significance for understanding PD pathophysiology. However, studies on the pathogenesis of the marmoset PD models can take a long time due to the long lifespans of the marmosets compared with rodents. Currently available genetic models do not completely induce appreciable neurodegeneration and PD phenotypes [35], whereas the neurotoxic models are used to damage the nigrostriatal pathway [10]. The marmoset model is a recognized model of PD using neurotoxins that induce the selective degeneration of nigrostriatal neurons [223637]. The most commonly used toxins are 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) and 6-hydroxydopamine (6-OHDA), which reproduce the pathological and behavioral changes of the human disease in rodents or NHPs. These models can be developed by the systemic or local administration of neurotoxins depending on the type of agent used and the species involved [9].
MPTP
MPTP was found to be a DA neurotoxin in the early 1980s when Langston et al. [38] described the occurrence of severe symptoms similar to PD in several young Californian intravenous drug users following the injection of an analogue of the narcotic meperidine that contained MPTP [39]. The identification of a specific neurotoxin, MPTP, that induces neural damage and other signs of PD in humans [39] has led to the development of valuable mammalian models, including sheep, dogs, guinea pigs, cats, mice, rats, and monkeys, for research on the pathophysiology, etiology, and pathogenesis of PD [4041]. The MPTP-primate model using baboons [42], macaques [4], velvet monkeys [43], squirrel monkeys [44] and marmosets [222324] remains the best model for preclinical evaluation of the efficacy of anti-parkinsonian therapy although most studies on PD research have been performed in mice due to a lack of resources and trained personnel for monkey models [1045].
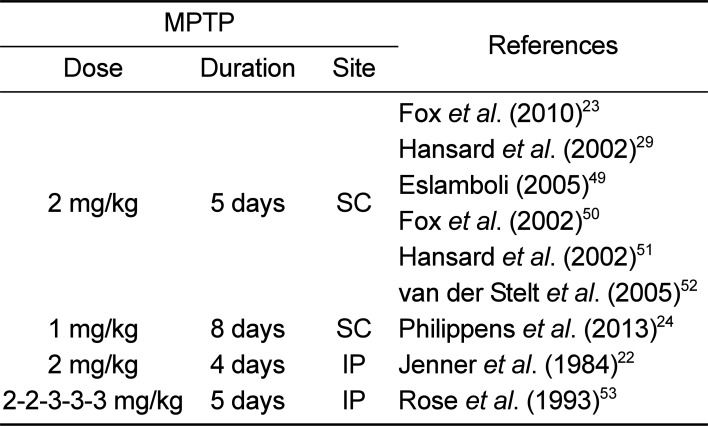
Since MPTP is a highly lipophilic compound, it can rapidly cross the blood-brain barrier (BBB). The mechanism underlying MPTP toxicity is the conversion of the MPTP by monoamine oxidase B into the final active toxin cation, 1-methyl-4-phenylpyridinium ion (MPP+), which can enter DA neurons in the substantia nigra pars compacta (SNpc) through the dopamine transporter (DAT). MPP+ induces neurotoxicity primarily by blocking mitochondrial complex I activity, leading to ATP depletion and increased oxidative stress [4647]. In general, a pathological limitation of the mouse MPTP model is the lack of LBs, which is the neuropathological landmark of PD [10], although a few reports have investigated the expression of a LB major constituent (á-synuclein) by modifying the MPTP treatment regimens [9]. Forno et al. [48] also described intraneuronal inclusions reminiscent of LBs in MPTP-injected monkeys. Several MPTP dosing regimens (Table 1) have been used in many studies [222324294950515253].
In comparison to other available PD models, the MPTP model does not require skilled stereotaxic surgery and is known to have the greatest similarity with human PD symptoms biochemically, anatomically and behaviorally [49]. However, in comparison to the 6-OHDA model, the MPTP model shows a dose-dependent risk of mortality from cardiotoxicity within 24 h of the first dose [54]. In addition, strict safety procedures are required with appropriate laboratory safety equipment since systemic MPTP treatment is a severe safety hazard to the personnel handling the animals [41]. Although MPTP itself is not directly harmful, its metabolite MPP+ is tremendously toxic. Since MPTP metabolites are excreted up to 3 days post administration [55], researchers should wear personal protection during all procedures involving MPTP including the preparation, injection, and 3-5 days post injection. Furthermore, all procedures involving MPTP should be conducted in a special room with a fume hood and enclosed cages under negative-pressure (Figure 1C) due to the MPTP aerosols generated from bedding, excreta and animal fur [41].
6-OHDA
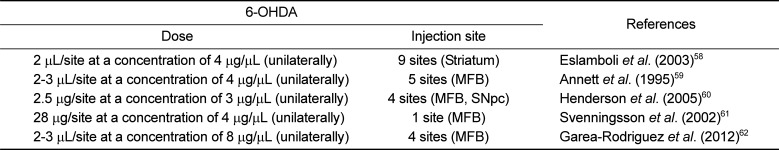
A catecholamine neurotoxin 6-OHDA is a hydroxylated analogue of DA with high affinity for DAT, which transports the toxin into DA neurons [9], and it can cause selective degeneration of sympathetic adrenergic nerve terminals [56]. The mechanism of 6-OHDA neurotoxicity has often been involved in the production of the reactive oxygen species, H2O2, resulting from the autoxidation of 6-OHDA [9]. It has also been reported that 6-OHDA can accumulate in the mitochondria, where it inhibits the mitochondrial respiratory enzymes (chain complex I), resulting in a damaging depletion of intracellular ATP and consequently cell death [957]. 6-OHDA has been used in rats, cats, guinea pigs, dogs, and monkeys [3140]. Since 6-OHDA does not cross the BBB, direct injection into the region of interest in the brain is required to target specific neurons, with several dosing regimens and different injection sites (Table 2) [5859606162]. Despite the difficulty in targeting small brain structures, the SN is usually targeted for 6-OHDA injection to create a more selective animal model of PD [60]. 6-OHDA lesions can also be made by targeting the striatum or the medial forebrain bundle (MFB). 6-OHDA injection into the striatum induces slow, progressive and partial damage of SNpc neurons over a period of up to 3 weeks, which is less marked than the effects of intra-MFB injection. In contrast, when injected into the MFB, which conveys the efferent fibers from nigral cell bodies to the striatum, 6-OHDA produces a rapid and massive degeneration in the nigrostriatal pathway. When injected into the nigra, a significant loss of striatal DA terminals is established within 2-3 days post-injection [910].
The administration of 6-OHDA is more complicated and time-consuming due to the necessity of stereotaxic surgery (Figure 2) and the difficulty in targeting small brain structures such as the SN or MFB. Additionally, 6-OHDA-treated animals fail to develop LBs, which are eosinophilic inclusions that contain ubiquitinated proteins such as α-synuclein [63]. Unilaterally lesioned animals have been used more often because bilateral injections can induce a far more severe phenotype with marked adipsia, aphagia, and high mortality. The major advantage of the unilateral 6-OHDA model is its feasibility for assessing a variety of different behaviors such as fine motor skills, sensorimotor neglect and body asymmetries. Also, the unilateral injection of 6-OHDA into one hemisphere in the animals leaves the unlesioned side as an internal control, hence, fewer animals are needed in experiments to analyze behavioral deficits [9]. Along with less postoperative care for animals, local 6-OHDA treatment can minimize the risk of inadvertent toxic exposure for researchers associated with systemic MPTP treatments [496465].
Rotenone
Rotenone is a flavonoid found in several plants and is a broad-spectrum pesticide used to kill insects. Like MPTP, it is highly lipophilic, so it readily crosses the BBB and diffuses into cells. Mitochondrial complex I inhibition, selective nigrostriatal neurodegeneration, and α-synuclein-positive cytoplasmic inclusions, which have been reported as key pathological features of clinical PD, were demonstrated in the rotenone model. Therefore, there has been interest in the PD model using rotenone [1066]. However, Betarbet et al. [66] indicated that rotenone displays systemic toxicity and subsequent high mortality rates (~30% of animals) regardless of the administration route. The main limitations of the rotenone model are variability in terms of the percentage of animals that develop a nigrostriatal DA lesion, the extent of the lesion, and the lesion distribution within the striatum [67]. Some animals can be resistant to rotenone (~50% of treated animals display neurodegeneration). This low success rate for the animal model of rotenone makes it necessary to use a larger number of animals at the start of any study. Nonetheless, the PD model involving systemic intravenous delivery of rotenone using osmotic pumps replicates many aspects of the pathology of human PD. Also, chronic intraperitoneal injection of rotenone causes locomotor impairment and neurobehavioral abnormalities characteristic of PD [6869]. Rotenone has been shown to induce DA neurodegeneration and Parkinson-like behavior through many pathogenic pathways including massive reactive oxygen species formation, proteosome activity inhibition, proteolytic stress, α-synuclein phosphorylation and aggregation and Lewy pathology [970]. Therefore, rotenone model can be used for the marmosets along with other classical PD models, MPTP and 6-OHDA.
Development of Parkinsonian Symptoms in the Marmosets
Clinical signs, artificial feeding, and intensive care
In order to identify the effects of parkinsonian agents or anti-parkinsonian treatments, body weight, complete blood count, serum biochemistry, gross and microscopic examination of tissues can be measured [71]. Following the induction of Parkinsonian symptoms, there is a critical need for daily management of the marmosets due to potential health threats (adipsia, aphagia, loss of body weight). Usually, the marmosets are allowed to have access to specially formulated pellets, fresh fruits, vegetables and water ad libitum. However, in order to maintain the body weight, welfare and survival of the animals displayed a stable level of akinesia following neurotoxin administration, artificial feedings consisting of a highly nutritious solution made from egg white, sucrose, infant formula, banana, multivitamins, and powdered marmoset pellets should be introduced on a daily basis by trained staffs until the marmosets are able to feed themselves. Also, the marmosets with very severe Parkinson's symptoms can be placed in an intensive care unit with tightly controlled temperature, humidity, and oxygen concentration (Figure 1D) until the marmosets are capable of maintaining food and water intake on their own. However, euthanasia is recommended for animals showing significant body weight loss (>20%) and anorexia.
Behavioral assessment
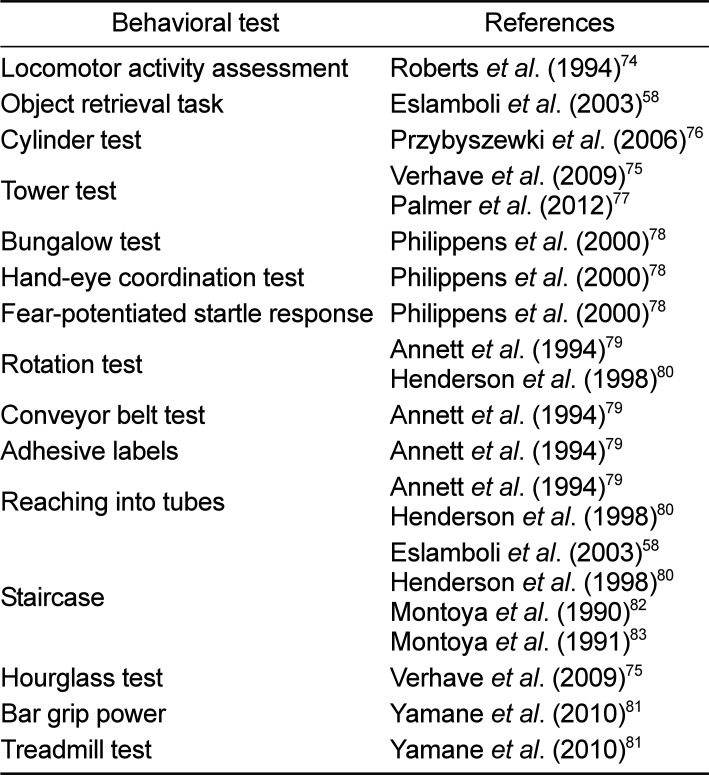
In order to cure Parkinsonian symptoms, highly reproducible animal models of PD should be developed to address all PD-related questions including pathological changes in the brain. First, the occurrence of typical PD symptoms can be recorded using a clinical rating scale (apathy, immobility, muscle rigidity, tremors and inadequate grooming) and for the purpose of scoring abnormal involuntary movements (facial behaviors, full body behaviors, and general severity of involuntary movements and incapacitation due to these movements) (Table 3) [36587273]. These clinical measurements can be performed in a double-blind manner by watching post hoc video-recordings of the animals accompanied by recording spontaneous locomotor activity in the lesioned animals [74]. Behavioral tests provide an external measure of PD-like pathology when assessing PD lesions in animals [71]. Evaluation of Parkinsonian features in marmosets can be conducted by a variety of behavioral tests ranging from simple food-retrieval task to a comprehensive neuropsychological assessment, including cylinder test, tower test (Figure 3), bungalow test, hand-eye coordination test, fear-potentiated startle response, rotation test, conveyor belt test, adhesive labels, reaching into tubes, staircase, hourglass test, bar grip power, and treadmill test (Table 4) [58757677787980818283].
Positron emission tomography (PET)
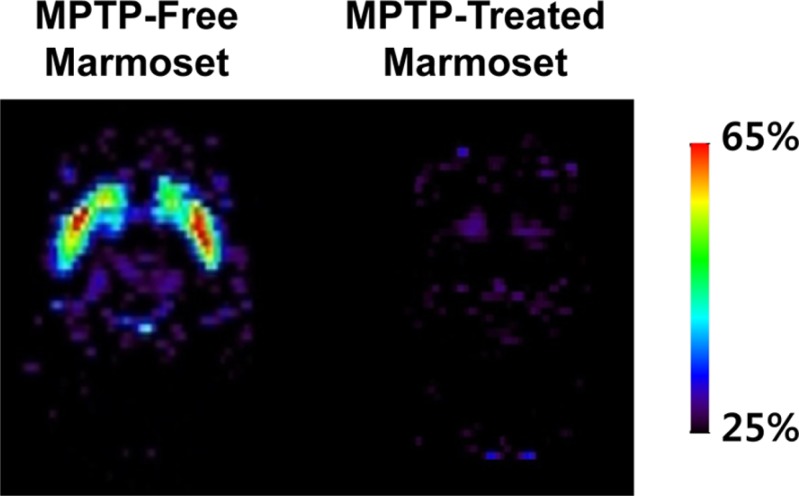
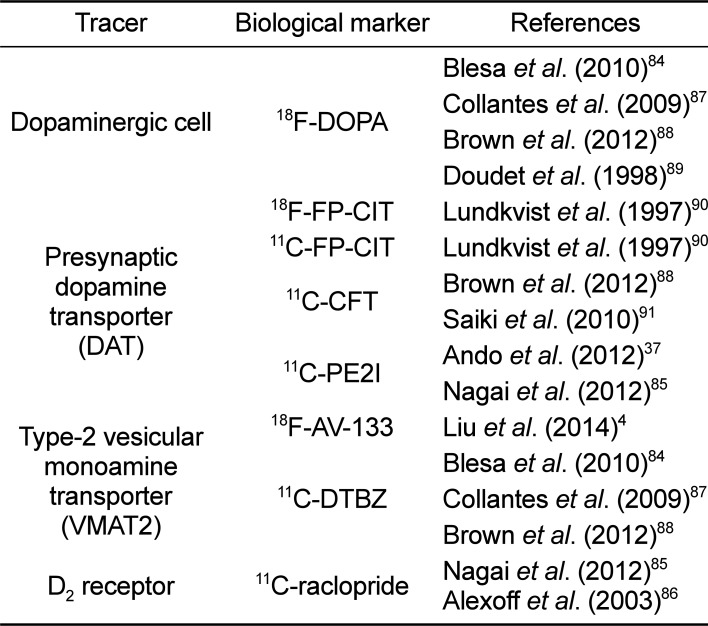
PET imaging is a relatively non-invasive neuroimaging technique that can be used to elucidate the functional changes associated with PD (Figure 4) and to provide diagnostic information by evaluating the integrity of the nigrostriatal DA system [48485]. In particular, a microPET scanner has been designed for the imaging of small-bodied animals, such as the marmoset. Under isoflurane anesthesia, the marmosets should be kept in a stereotaxic unit throughout the scanning process. Tracer compounds labeled with positron-emitting radioisotopes, including 18F-DOPA, 18F-FP-CIT, 11C-FP-CIT, 11C-CFT, 11C-PE2I, 18F-AV-133, 11C-DTBZ, and 11C-raclopride, should be administered to image and measure biochemical processes in vivo (Table 5) [4378485868788899091].
PET studies with 18F-DOPA [84878889] have been used to examine the dysfunction of the nigrostriatal DA system. 11C-raclopride [8586] specifically binds to dopamine D2 receptors. Also, 18F-AV-133 [4] and 11C-DTBZ [848788] target the vesicular monoamine transporter type 2 (VMAT2), which is a vesicular membrane protein that transports monoamines (including DA, norepinephrine, and serotonin) into synaptic vesicles. DATs are located on DA nerve terminals. The DAT-selective ligands 18F-FP-CIT [90], 11C-FP-CIT [90], 11C-CFT [8891], and 11C-PE2I [3785] have been used as a marker of DA nerve degeneration. When DA nerve terminals in the nigrostriatal system degenerate in PD, DAT is not available to bind to these ligands.
Magnetic resonance imaging (MRI)
The rapid advancement in functional and structural brain imaging has made it feasible to quantify specific changes in brain function within the larger perspective of whole-brain systems [92]. MRI is far more widely available than PET and is the most commonly used system for anatomical imaging of the entire brain in vivo. It can non-invasively acquire functional images showing openings in the BBB, the presence of lesions caused by the Parkinsonian agent [9394]. Marmosets, which are a suitable species for PD research, have been increasingly studied with MRI. Unlike larger monkeys, MRI scans are not needed for the marmoset because a standard frame-based stereotactic apparatus fitted with a small primate head-holder for a variety of neurosurgical procedures is sufficient to perform reproducible intracerebral surgeries [4995]. MR spectroscopy can also be utilized as a non-invasive tool to assess in vivo dynamic changes associated with neurodegeneration and anti-Parkinsonian treatments by measuring the presence and concentration of certain metabolites, especially in MR spectra acquired at high magnetic field strengths and with short echo times [9697].
Biomarkers for PD
Biomarkers are needed as indicators for the diagnosis and monitoring of disease progression in PD. DJ-1, a redox-sensitive molecular chaperone protein, appears to be linked to PD with oxidative stress in the mitochondria and nucleus [9]. Also, a pathological sign of PD is the presence of intracellular proteinaceous inclusions, LBs, which are composed mainly of α-synuclein [37]. Consequently, DJ-1 and α-synuclein are considered helpful diagnostic markers for PD in human cerebrospinal fluid as well as in plasma or serum [98] although the use of plasma DJ-1 and α-synuclein as biomarkers is controversial [99]. As tyrosine hydroxylase (TH) catalyses the formation of L-DOPA associated with the biosynthesis of DA [100], immunostaining for TH-positive neurons and DAT have been used to assess nigral DA neuron death after MPTP administration [45]. It has also been reported that significant increases in the pro-inflammatory cytokines IFN-γ and TNF-α in the blood serum samples of MPTP-treated monkeys were observed with ELISA measurements, indicating a critical role in stimulating and maintaining glial cell activation in the SNpc as well as contributions to DA neuronal degeneration and motor impairment Parkinsonism [101].
Animal Models for Assessing Novel Treatments
A number of anti-Parkinsonian drugs, including DA drugs (apomorphine, pramipexole, ropinirole, pergolide, bromocriptine and cabergoline), muscarinic antagonists (benztropine and trihexyphenidyl), monoamine oxidase-B inhibitors (selegiline and rasagiline), and catechol-O-methyltransferase inhibitor (tolcapone), are in current clinical use for PD. These drugs have shown different levels of predictive validity for efficacy depending on the animal models, which reflect different degrees of the pathology and biochemical changes associated with PD [63]. To determine the appropriateness of the model for the discovery of novel treatment for PD symptoms, the similarity of the model with the human disease is especially important. On a basis of the strengths and weaknesses of the respective available models, the researchers should select optimal experimental animal models of PD depending on different target mechanisms to find potential drugs for therapy. In this respect, primate (marmoset) models are particularly useful. More importantly, the fact that the MPTP model possesses the greatest similarity to the clinical features of human PD disease such as tremors, rigidity, akinesia, and postural instability can lead researchers to consider it the most clinically-relevant model for late phase preclinical assessment of a novel treatment [3749]. However, the 6-OHDA model also has an advantage associated with the unilateral lesion to dissociate different symptoms of DA loss [63]. With the growth of biotechnology-derived products, stem cell therapy and gene therapy can be considered potential and novel therapies that provide a more permanent remedy than current drug treatments. Takagi et al. [102] reported that the transplantation of DA neurons generated from monkey ES cells resulted in attenuation of MPTP-induced neurological symptoms in the primate MPTP model for PD as evidenced by behavioral studies and functional imaging. In addition, Kikuchi et al. [103] demonstrated that human induced pluripotent stem cell-derived neural progenitor cells survived for six months as DA neurons in the brain of the primate MPTP model for PD. Lentiviral delivery of glial cell line-derived neurotrophic factor into the striatum and SN has been shown to reverse functional deficits and to prevent nigrostriatal degeneration in the primate MPTP model for PD [104].
Conclusion
In general, research laboratories and pharmaceutical/biotechnology companies follow a sequence of steps for the research and development of treatments related to a variety of important human diseases. An important stage in this process includes the determination of appropriate animal models for predicting the effectiveness of the treatment strategies in clinical trials. Although animal models may not sufficiently reflect the features of the human disease, they can be used to analyze particular aspects and pathogenic mechanisms of the disorder which cannot be fully studied using simple in vitro models. Interest in NHPs is increasing since they are the only relevant species that show high similarity to humans for preclinical assessment prior to clinical trial. At the preclinical stage, use of the marmoset as an experimental model offers several considerable advantages such as smaller size, early sexual maturation, and rapid breeding in pharmaceutical product development, resulting in reduced test material requirements and earlier assessment of product candidates in adults. In particular, the marmoset can be used as an appropriate PD model that reflects various aspects of the human disease as well as an experimental subject for adequate safety or efficacy assessments of the therapeutic treatment, especially with stem cell therapy and gene therapy for PD. Currently, the significant progress in the production of transgenic marmoset models for PD is being made. However, the neurotoxic models are relatively easy to induce parkinsonism in NHPs including the marmosets although the animals with severe symptoms following the systemic or local administration of the neurotoxins need the intensive care with artificial feedings. By providing an overview of the models, methods, and animal care procedures associated with PD research, this article may help researchers with the selection of appropriate animal PD models depending on the specific objectives and aims of their study.




 XML Download
XML Download