

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Hypertension is a major risk factor for hypertensive heart disease, coronary artery disease, stroke, aortic aneurysm, and peripheral arterial disease [1,2]. The induction of hypertension is correlated with dysfunction of the vascular endothelium, which can greatly influence vascular tone and structure. In the case of human hypertension, vascular dysfunction was observed in peripheral and coronary macrocirculation and microcirculation, as well as in renal circulation [3]. In endothelial cells, hypertension can be regulated by the release of various contracting factors, including NO, prostanoids, oxygen free radicals and endothelin-1 [4,5]. Thus, prevention and improvement of endothelial dysfunction can provide a strong advantage in the therapeutic strategy of patients with cardiovascular risk factors.
Accumulated evidence has recently indicated that antioxidant capacity is associated with reduced risk of cardiovascular disease. Some antioxidant substances that improve endothelium dysfunction can be expected to decrease the incidence of cardiovascular disease. Aqueous extracts obtained from Thymus serpyllum (TE) with high antioxidant capacity induced a significant decrease in systolic and diastolic blood pressure and total peripheral resistance in SHR, without affecting these parameters in WKY rats [6]. Moreover, a similar response was observed after long-term administration of sesamin, one of the major lignans in sesame seed with antioxidative and antihypertensive activities. SHR treated with sesamin showed improvement of arterial function through increased eNOS expression and decreased p22phox and p47phox expression [7]. Silk fibroin hydrolysate (SFH) exhibited a blood pressure-lowering effect in SHR accompanied with a decline in endothelin and NO concentration [8,9]. However, the calcium antagonist nifedipine did not induce a significant decrease of systolic blood pressure in stroke-prone spontaneously hypertensive rats (SHRSP), even though they have the capability for upregulation of Cu/ZnSOD expression and activity [10].
Several previous approaches have shown the novel functions of RLP on improvement of human disease after its preparation using a steaming process [11]. Aqueous extract of RLP (AEtRLP) treatment induced down-regulation of glucose, stimulation of insulin secretion, activation of the insulin receptor signaling pathway and Glucose transporter 2 (GLUT2) and 3 expression in INS cells and streptozotocin-induced diabetic mice [11,12]. Moreover, OLETF rats treated with AEtRLP showed dramatic alterations in insulin secretion and fatty acid metabolism. The triglyceride and low density lipoprotein (LDL) concentration and fatty liver formation were reduced dramatically in the AEtRLP treated group when compared to the vehicle treated group [13]. Moreover, AEtRLP stimulated the secretion of nerve growth factor (NGF) from B35 neuronal cells as well as the brains of NSE/hAPPsw transgenic mice expressing Alzheimer's disease [14,15]. Therefore, additional studies are needed to identify the novel functions of RLP to determine if they have the potential for use as therapeutic strategies for the treatment of chronic diseases showing vascular dysfunction.
In this study, we investigated the beneficial effects induced by repeated administration of EEtRLP using SHR. The results provide novel data indicating that EEtRLP will be useful for improvement of vascular dysfunction in hypertension response.
Materials and Methods
Preparation of EEtRLP and RLP
RLP was prepared from the roots of LP using the methods described in previous studies [16]. Briefly, fresh roots of LP were collected from plantations in the Miryang area (Korea) and sufficiently dried in a hot-air drying machine (JSR, Seoul, Korea) at 60℃. Voucher specimens of LP (WPC-11-010) were deposited in the Functional Materials Bank of the PNU-Wellbeing RIS Center at Pusan National University. To produce RLP9 at nine different steaming frequencies, 200 g of dry roots were steamed at 99℃ for 3 h and then air-dried at 70℃ for 24 h a total of nine times (Figure 1A). Final produced RLP9 roots were reduced to powder using an electric blender, after which ethanol extracts (EEt) were purified for 18 h at 25℃ using a shaking incubator (JSR Research Com., Kongju, Korea) following the addition of 200 mL of 70% ethanol into the RLP9 powder (10 g). In addition, a solution of the extract was concentrated to dry pellets in a rotary evaporator (EYELA, Tokyo, Japan) and stored at -80℃ until needed.
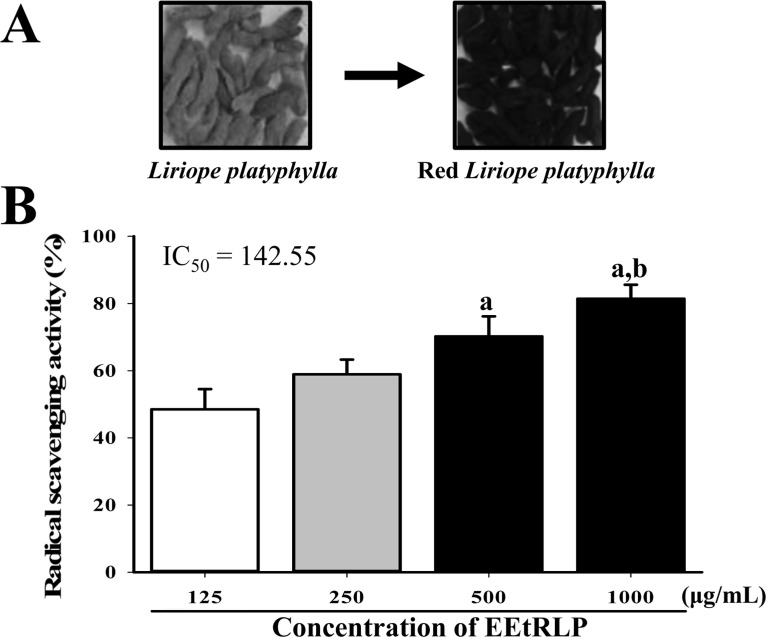 | Figure 1Morphology of RLP and DPPH scavenging activity of EEtRLP. (A) RLP was prepared from roots of LP by steaming at 99℃ for 3 h followed by air-drying at 70℃ for 24 h nine times. During this process, their color gradually changed from yellow to dark. (B) DPPH scavenging activity of EEtRLP was measured at four different concentrations. The inhibitory concentration (IC50) was calculated from these results. Data shown are the means±SD (n=5). a, P<0.05 compared to the 125 µg/mL. b, P<0.05 compared to 250 µg/mL.
|
Analysis of total flavonoid and phenolics
Total phenolics were measured by the Folin-Ciocalteu method, with slight modification [17]. Briefly, 1 mL of EEtRLP solution was mixed with 5mL of Folin-Ciocalteu reagent (Sigma-Aldrich Co., St. Louis, MO, USA) and then incubated at room temperature for 5 min. The mixture was subsequently added to 15 mL of 20% Na2CO3 and vortexed for 30 sec, after which the absorbance was repeatedly measured at 765 nm using a Versa-max plate reader (Molecular Devices, Sunnyvale, CA, USA). A standard calibration curve was made using different concentrations of gallic acid (Sigma-Aldrich Co.), and the concentration of total phenolic contents in EEtRLP was presented as mg gallic acid equivalent of extract.
The total flavonoid contents were measured as previously described [18]. Briefly, 200 µL of several different concentrations of EEtRLP were mixed with 60 µL of 5% NaNO2 (Sigma-Aldrich Co.) and 60 µL of 10% AlCl3 (Sigma-Aldrich Co.). Following incubation at 25℃ for 5 min, the mixture was added to 400 µL of 1M NaOH and the absorbance was repeatedly measured at 510 nm using a Versa-max plate reader (Molecular Devices). A standard calibration curve was then made using different concentrations of catechin (Sigma-Aldrich Co.). The concentration of flavonoid contents in EEtRLP was presented as mg catechin equivalent of extract.
Free radical scavenging activity
The scavenging activity of DPPH radical was measured as previously described [19]. Briefly, each sample (100 mL) in four different concentrations of EEtRLPs (125, 250, 500 and 1,000 µg/mL) was mixed with 100 mL of 0.1 mM DPPH (Sigma-Aldrich Co.) in 95% ethanol solution or 100 mL of 95% ethanol solution and then incubated for 30 min at room temperature. After the reaction time, the absorbance of the reaction mixture was measured at 517 nm using a Versa-max plate reader (Molecular Devices, Sunnyvale, CA, USA). DPPH radical scavenging activity of the EEtRLP was expressed as the percent decrease in absorbance relative to the control. The IC50 value is defined as the concentration of substrate that causes a 50% loss in DPPH activity.
Design of animal experiment
The animal protocols used in this study were reviewed and approved based on the ethical and scientific care procedures of the Pusan National University-Institutional Animal Care and Use Committee (PNU-IACUC; Approval Number PNU-2012-0134). The 16-week-old WKY and SHR used in this study were purchased from Central Lab Animal Inc. (Seoul, Korea). All rats were provided with ad libitum access to a standard irradiated chow diet (Samtako, Osan, Korea) and water throughout the 14-day feeding study. This diet was composed of moisture (12.5%), crude protein (25.43%), crude fat (6.06%), crude fiber (3.9%), crude ash (5.31%), calcium (1.14%), and phosphorus (0.99%), as well as corn (546 g/kg), vegetable protein (316 g/kg), fish meal (34 g/kg), beet pulp (30 g/kg), animal fat and oil (36 g/kg), lysine (2 g/kg), choline bitartrate (2 g/kg), 7.17% methionine solution (2 g/kg), limestone (10 g/kg), calcium/phosphate supplement (13 g/kg), salt (5 g/kg), mineral mix (2 g/kg), and vitamin mix (2 g/kg). During the experiment, rats were maintained in a specific pathogen-free state under a strict light cycle (lights on at 08:00 hours and off at 20:00 hours) at 23±2℃ and 50±10% relative humidity. Both types of rats were handled in the Pusan National University-Laboratory Animal Resources Center accredited by the Korea Food and Drug Administration (KFDA) (Accredited Unit Number-000231) and AAALAC International according to the National Institutes of Health guidelines (Accredited Unit Number-001525).
Sixteen-week-old SHR (n=12) were randomly divided into three groups: (1) SHR treated with phosphate buffer saline (PBS) (vehicle treated group), (2) SHR treated with 100 mg/kg of EEtRLP (EEtRLP100 treated group), (3) SHR treated with 500mg/kg of EEtRLP (EEtRLP500 treated group). Another four WKY rats were used as a control group. Two concentrations (100 and 500 mg/kg) of EEtRLP were separately dissolved in PBS and administered to rats orally every day for a period of 14 days. At the end of the period, all rats were euthanized with carbon dioxide gas and blood and tissue samples were collected.
Measurement of blood pressure
The systolic blood pressure was measured with a non-invasive blood pressure measurement system using the tail-cuff method (Non-invasive Blood Pressure Analyzer 179, NTC Technology Inc. Rancho Cordova, CA, USA). The systolic blood pressure (SBP), diastolic blood pressure (DBP), mean blood pressure (MBP) and heart rate (HR) were measured as previously described [20]. The blood pressure of the rats was measured, while the animals were awake without harming the animal. Blood pressure values were recorded by a pressure transducer and data acquisition system (NTC Inc. Life Science Instruments). Before measuring the blood pressure, all rats were allowed to stabilize for at least 30-40 min. Two concentrations of EEtRLP were administrated to rats daily for 14 days. At 5 h after final treatment, the blood pressure of all rats was measured in triplicate. Rats were allowed to habituate to this procedure for at least 1 week prior to the experiments.
Histological analysis
The thoracic aorta was removed from rats of subset groups, fixed in 10% formalin, embedded in paraffin wax, routinely processed, and then sectioned into 4 µm thick slices. Next, the aorta sections were stained with hematoxylin and eosin, after which they were examined by light microscopy at 400× magnification to determine the cellular morphology. The thickness of the individual smooth muscle cell layers was measured using the Leica Application Suite (Leica Microsystems, Wetzlar, Germany).
Western blotting
Thoracic aorta collected from a subset of the groups (n=4 per group) was homogenized using a PRO-PREP™ Solution Kit (iNtRON Biotechnology, Sungnam, Korea) supplemented with half of a protein inhibitor cocktail tablet (Roche, Penzberg, Germany). After homogenization, samples were centrifuged at 10,000×g for 10 min. The prepared proteins were then electrophoresed through a 10% SDS-PAGE gel, after which they were transferred to nitrocellulose membrane (Amersham Biosciences, Corston, UK) for 2 h at 45 V in transfer buffer (25 mM Trizma-base, 192 mM glycine, and 20% methanol). The efficiency of the transfer and equal protein loading were evaluated by staining the membrane with Amido Black Staining Solution (Sigma-Aldrich Co.) and the gel with Coomassie Blue. Appropriate dilutions of primary antibodies, anti-ACE antibody (R&D systems, Minneapolis, MN, USA), anti-ACE2 antibody (R&D systems), anti-eNOS antibody (Abcam, Cambridge, UK) and anti-β-actin (Sigma-Aldrich Co.) were added to the membranes and allowed to hybridize overnight at 4℃. After the antibodies were removed, the membrane was washed three times in a solution composed of 10 mM Trizma-base (pH 7.6), 150 mM NaCl, and 0.05% Tween-20 for 10 min each. The membrane was subsequently incubated with horseradish peroxidase-conjugated anti-secondary antibody for 1 h at room temperature, after which it was washed again as described above and developed using an enhanced chemiluminescence detection system (Amersham Bioscience). The results were quantified using the Image Analyzer System (Eastman Kodak 2000MM, NY, USA) and expressed as the fold-increase over control values.
Detection of aldosterone concentration
The concentration of aldosterone in serum was measured by radioimmunoassay with a Coated A count Aldosterone kit (Siemens, CA, USA) according to the manufacturer's suggestions. First, serum (200 µL) collected from whole blood and 125I-aldosterone were added into an aldosterone Ab-coated tube and vortexed briefly and gently. Following incubation for 18 h at room temperature, all visible moisture was decanted thoroughly from the aldosterone Ab-coated tube, after which samples were allowed to drain for 2 or 3 min. Finally, the radioactivity of 125I-aldosterone was measured with γ-counter (Quantum, Parkard, USA) and the aldosterone concentration was calculated using the following equation:
Measurement of NOx concentration
The plasma NOx concentration was determined using a Total Nitric Oxide and Nitrate/Nitrite Assay Kit (R&D systems) according to the methods suggested by the manufacturer. To measure the concentration of endogenous nitrite present in the sample, 50 µL of each sample and standard were incubated for 10 min at 25℃ in a 250 µL incubation mixture composed of reaction diluent and Griess reagent I and II. The optical density of each well was determined spectrophotometrically at 540 nm. To determine the nitrate concentration in the sample, 50 µL of each sample and standard were incubated for 30 min at 37℃ in a 150 µL incubation mixture composed of reaction diluent, NADH and nitrate reductase. This reaction was initiated by the addition of nitrate reductase to convert nitrate to nitrite. Finally, a chain of reaction was terminated by the addition of 50 µL of Griess reagent I and II for 10 min at room temperature. After centrifugation at 15,000 rpm for 5 min, the absorbance of supernatants from each mixture was determined spectrophotometrically at 540 nm. The level of NOx was determined by adding the nitrite concentration and the nitrate concentration.
Determination of MDA levels
The MDA level was assayed using a Lipid Peroxidation (MDA) Assay Kit (Sigma-Aldrich Co.) according to the manufacturer's protocols. Briefly, the plasma collected from each rat was mixed with 42 mM H2SO4, after which phosphotungstic acid solution was added. Sample solution was incubated at room temperature for 5 min and then centrifuged at 13,000×g for 3 min. The pellet was subsequently mixed with deionized water containing butylhydroxytoluene (BHT). The sample or standards and TBA solution (70 mM thiobarbituric acid, 5.0M glacial acetic acid) were incubated in a microcentrifuge tube at 95℃ for 60 min, then cooled to room temperature in an ice bath for 10 min, after which the reaction absorbance at 532 nm was read using a Versamax plate reader (Molecular Devices).
Measurement of SOD activity
The SOD activity in the serum collected from a subset of the groups was detected using a calorimetric assay and the reagents in the SOD Assay kit (Dojindo Molecular Technologies, Tokyo, Japan). To measure the SOD activity, the serum sample was diluted with dilution buffer or saline as follows: 1, 1/5, 1/52, 1/53, 1/54, 1/55, 1/56. Sample solution (25 µL) was then aliquoted into the wells of a 96-well plate for each blank or sample, after which 200 µL of WST working solution was added. In addition, an enzyme working solution (20 µL) was added to each sample well and mixed thoroughly. The enzyme reaction was then induced by incubating the mixture plate at 37℃ for 20 min, after which the absorbance was measured using a spectrophotometer at 450 nm. Finally, the SOD activity was calculated directly using the following equation: SOD activity (inhibition rate %)=[Ablank 1-Ablank 3)-(Asample-Ablank 2)]/(Ablank 1-Ablank 3)×100 (Ablank 1, absorbance of blank 1; Ablank 2, absorbance of blank 2; Ablank 3, absorbance of blank 3; Asample, absorbance of sample).
Detection of ion concentration
Following administration of the final treatment, all rats in each group were fasted for 8 h, after which blood was collected from the abdominal veins of rats and incubated for 30 min at room temperature. Serum was then obtained by centrifugation of whole blood. The concentrations of three ions (Na, K and Cl) in serum were assayed using an Automatic Serum Analyzer (HITACHI 747, Tokyo, Japan). All assays were conducted in duplicate using fresh serum.
Statistical analysis
One-way ANOVA was used to determine if significant differences existed between the WKY group and SHR group (SPSS for Windows, Release 10.10, Standard Version, Chicago, IL, USA). Additionally, differences in response between the vehicle treated group and EEtRLP treated group were evaluated by a post hoc test (SPSS for Windows, Release 10.10, Standard Version) of the variance and significance levels. All values were expressed as the means±SD. P values<0.05 were considered significant.
Go to : 
Results
Bioactive components and free radical scavenging activity of EEtRLP
As shown Table 1, EEtRLP contained high concentrations of two important antioxidants, flavonoids (343.0 µg/mL) and phenolics (40.4 µg/mL). Also, DPPH scavenging activity was dependent on the EEtRLP concentration, which ranged from 125 to 1,000 µg/mL. Significantly enhanced activity was detected in the 500 mg/mL and 1,000 mg/mL treated group compared to the 125 mg/mL treated group, while the highest level of activity was observed in the 1,000 mg/mL treated group (Figure 1B). These findings suggest that EEtRLP have high anti-oxidative activity.
Effects of EEtRLP on blood pressure and aortic wall thickness of the SHR
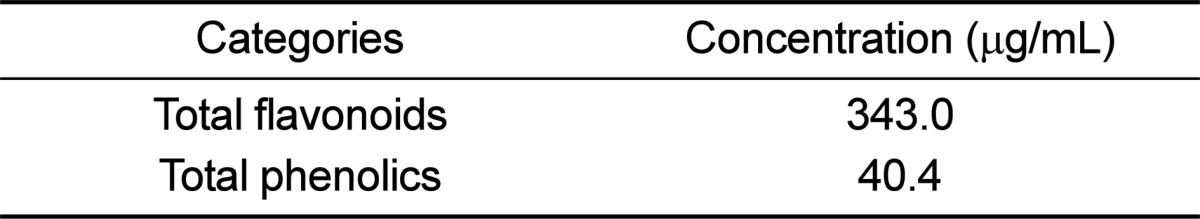
To determine if repeated administration of EEtRLP can relieve hypertension symptoms, alterations in systolic blood pressure and abdominal aortic wall thickness were measured in SHR after EEtRLP treatment for 2 weeks. High blood pressure was only detected in the SHR group compared to WKY rats. However, blood pressure levels were not recovered, but were instead maintained a constant level in the EEtRLP treated group (Figure 2A). Moreover, alterations in the abdominal aortic wall thickness were very similar to those in systolic blood pressure. Specifically, the thickness was significantly greater in SHR than WKY under the no treatment conditions. After EEtRLP treatment for 2 weeks, their levels were not recovered to those of the WKY (Figure 2B). Taken together, these findings suggest that EEtRLP administration for 2 weeks did not induce any significant alteration in aortic wall thickness or blood pressure in SHR.
 | Figure 2EEtRLP effect on systolic blood pressure and the histological structure of thoracic aorta. (A) SHR and WKY rats were treated with two different doses of EEtRLP (100 and 500 mg/kg) for 2 weeks. Prior to measurement of blood pressure, all rats were allowed to habituate to the procedure for 5 h after final treatment. (B) The slide sections of the thoracic aorta were stained with hematoxylin & eosin and then observed at 400× magnification. The thickness of tunica adventitia and smooth muscle were measured using the Leica Application Suite. Data shown are the means±SD (n=5). a, P<0.05 compared to the WKY rats.
|
Effects of EEtRLP on ACE and ACE2 expression in the aorta of SHR
We also investigated whether the effects of EEtRLP on vascular dysfunctions occurred despite the absence of significant symptoms of hypertension in SHR rats being detected in the EEtRLP treated group. To accomplish this, the level of hypertension regulatory proteins was measured in the aorta of SHR using specific antibodies. The expression level of ACE was about two times higher in the vehicle treated group than in WKY rats. Following EEtRLP treatment, their level was further increased by 21-63% compared to the vehicle treated group. However, the expression of ACE2 differed from that of ACE. The vehicle treated group showed a much lower level of ACE2 expression than WKY, while the EEtRLP treated group showed 20-22% enhancement compared to the vehicle treated group (Figure 3). These results suggest that EEtRLP could stimulate increased ACE and ACE2 expression in the endothelial cells of the aorta in SHR.
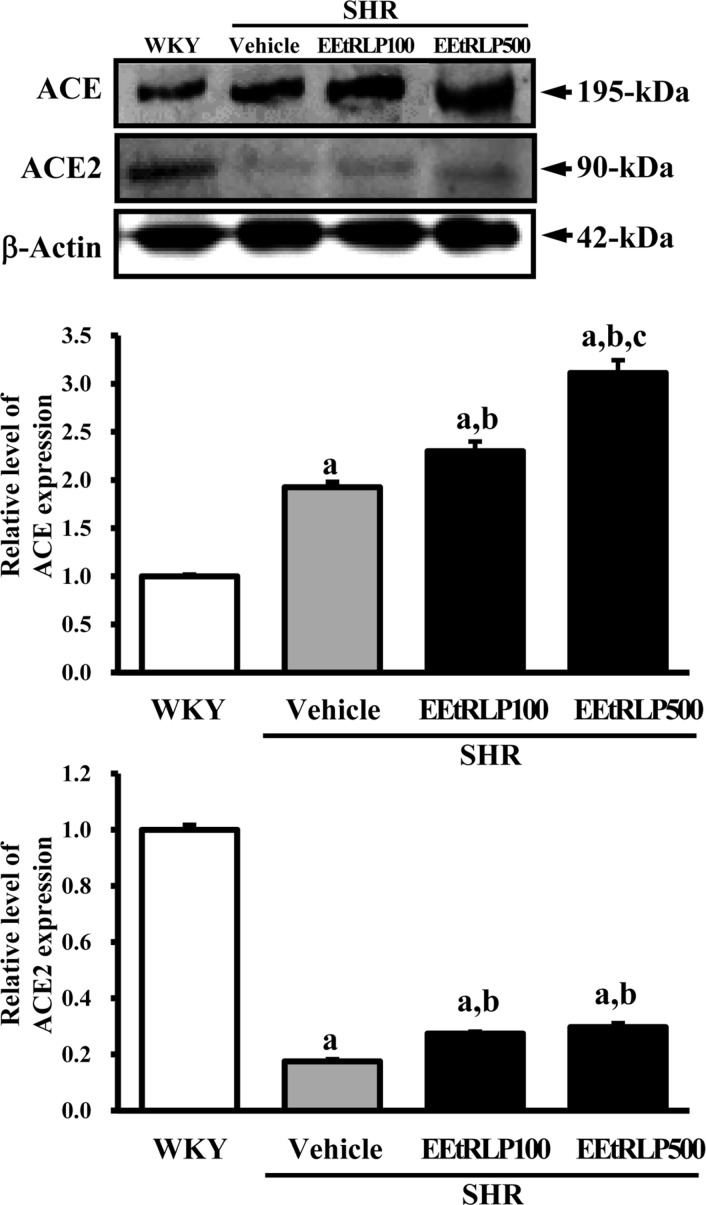 | Figure 3ACE and ACE2 expression in thorasic aorta. After detection of specific bands with two primary antibodies, the intensity of each band was determined using an imaging densitometer and the relative level of each protein was calculated based on the intensity of actin protein as an endogenous control. Data represent the mean±SD of three replicates. a, P<0.05 compared to WKY rats. b, P<0.05 compared to the vehicle-treated group within SHR.
|
Effects of EEtRLP on the concentration of aldosterone and ions in the serum of SHR
Aldosterone secreted from kidneys could contribute to the regulatory mechanism responsible for differences in some ion concentrations following treatment with various concentrations of anti-hypertensive agents [21]. Therefore, to determine if EEtRLP treatment led to alterations in the aldosterone regulatory system, the concentrations of aldosterone and related ions were measured in the serum of SHR after treatment for 2 weeks. A significant decrease in aldosterone concentration was observed in the EEtRLP treated group, although higher concentrations of aldosterone were detected in the vehicle treated group compared to WKY rats. Specifically, the concentration of aldosterone was 90.7% lower in the EEtRLP500 treated group than the vehicle treated group, while it was 75.6% lower in the EEtRLP100 treated group (Figure 4A). To determine if regulation of the aldosterone concentration was accompanied by the induced change in the related ions concentration, the levels of K, Na and Cl ions in serum were measured after EEtRLP treatment. Significant changes were only observed for the K concentration of the EEtRLP treated group, with a regulation pattern very similar to that of aldosterone being observed. The K concentration of the EEtRLP treated group was lower than that of the vehicle treated group, while that of the vehicle treated group was higher than the WKY group (Figure 4B). However, the concentration of Na and Cl remained constant in all experimental groups (Figure 4C and D). Overall, these results showed that EEtRLP treatment may stimulate a recovered serum K concentration through regulation of aldosterone, but the Na and Cl concentrations were maintained.
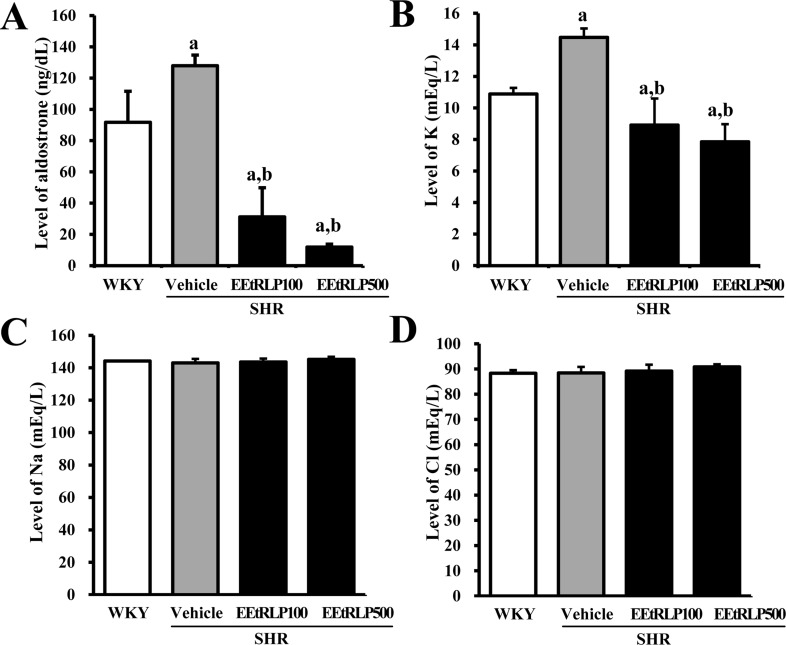 | Figure 4Aldosterone and ions concentration. (A) The concentration of aldosterone was measured in the serum of a subset group as described in the materials and methods. This method can generally detect a broad range of aldosterone concentrations (from 1,200 to 25 pg/mL). (B-D) The concentrations of three ions (K, Na and Cl) were measured in serum of a subset group using an Automatic Serum Aanalyzer. Data represent the mean±SD from three replicates. a, P<0.05 compared to WKY rats. b, P<0.05 compared to the vehicle-treated group within SHR.
|
Effects of EEtRLP on the expression of eNOS in the aorta of SHR
To investigate whether EEtRLP administration can induce increased anti-oxidative enzyme expression, the level of eNOS expression was measured in the aorta of SHR after EEtRLP treatment. As shown in Figure 5A, treatment with two doses of EEtRLP was found to induce an increase in eNOS expression in the SHR group, while their level was much lower in the vehicle treated SHR group than in WKY (Figure 5A). Taken together, these data suggest that EEtRLP treatment for 2 weeks may induce enhancement of eNOS expression in the aorta of SHR.
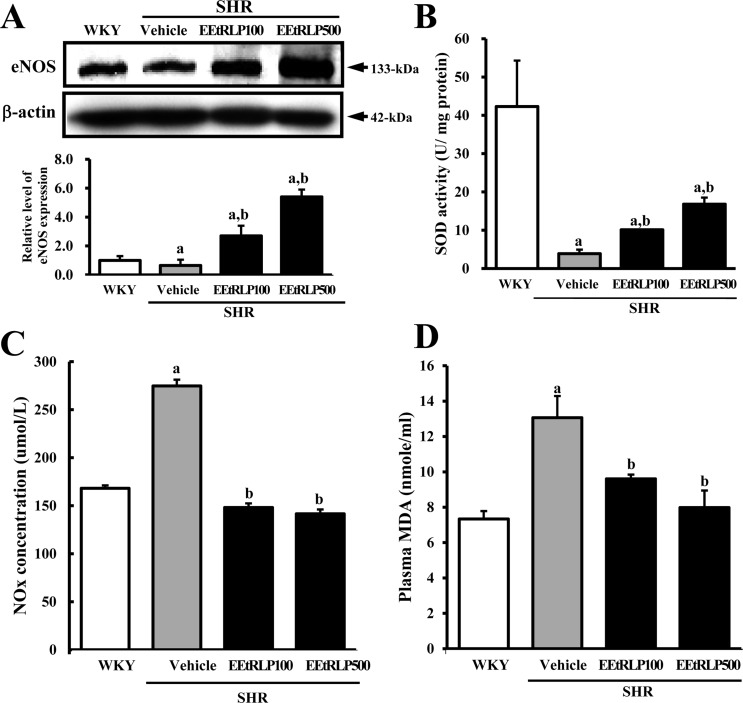 | Figure 5Alteration of antioxidant related factors. (A) After detection of specific bands with anti-eNOS antibodies, the intensity of each band was determined using an imaging densitometer and the relative level of each protein was calculated based on the intensity of actin protein as an endogenous control. (B) SOD activity, (C) NOx concentration and (D) MDA level were detected in the serum of a subset group using the appropriate kit. Data represent the mean±SD from three replicates. a, P<0.05 compared to the WKY rats. b, P<0.05 compared to the vehicle-treated group within SHR.
|
Effects of EEtRLP on SOD activity and MDA and NOx concentration in the serum of SHR
To determine if EEtRLP treatment could induce an antioxidant condition, the level of related factors including SOD activity and MDA and NOx concentration were measured in the serum of SHR. SOD activity was significantly decreased in the vehicle treated group when compared to WKY rats. Following EEtRLP treatment, SOD activity gradually increased to that of WKY rats (Figure 5B); however, an opposite pattern of SOD activity was observed for the concentration of MDA and NOx. As shown in Figure 5C and D, higher levels of MDA and NOx were detected in the vehicle treated SHR than WKY rats, whereas a greater decrease of MDA and NOx concentration were detected in the EEtRLP treated group. Taken together, these findings indicate that EEtRLP treatment can induce an increase in antioxidant condition, which might contribute to regulation of the renin-angiotensin system.
Go to :
Discussion
The antioxidative compounds and extracts of medicinal plants have been shown to reduce the risk of cardiovascular disease, partly due to improvement of vascular endothelial dysfunction [7]. However, further studies are needed to provide additional evidence regarding the correlation between antioxidative capacity and endothelium function. Therefore, we characterized the antioxidative capacity of EEtRLP and investigated the therapeutic mechanism of EEtRLP in SHR after administration for 2 weeks. The results presented herein provide evidence that EEtRLP treatment may induce improved endothelial dysfunction via upregulation of ACE expression, decreased aldosterone and K concentration, and enhanced eNOS expression and SOD activity.
Many investigations of the capacity of antioxidant compounds isolated from natural products have been conducted using DPPH and NO scavenging activity assays [22]. Sesamin, which has antihyperlipidemic, antioxidative and antihypertensive pharmacological properties in several animal models, was found to have an IC50 value of 2,508.63 µg/mL upon DPPH assay [23]. The aqueous extract obtained from Thymus serpyllum L. (wild thym: TE) showed an IC50 value of 122.36 µg/mL for NO scavenging activity, while TE protected against hypertension in an experimental model [6]. Moreover, the IC50 value of several compounds isolated from Picrasma quassiodes (D. Don) Benn (PQB) with antihypertensive effects had NO scavenging activity that ranged from 2.87 to 14.58 µg/mL [24,25,26]. In this study, the IC50 value of EEtRLP was very similar to that of TE, although the target molecule for the assay differed in both studies. However, the IC50 value of EEtRLP containing many kinds of unknown compounds was 10 times higher than that of the single compound.
A significant improvement in systolic blood pressure was observed in the SHR model following treatment with various antioxidants. Bolus injection of TE induced a significant decrease (39.2%) of systolic and diastolic blood pressure and total peripheral resistance in SHR [6]. Moreover, a similar decrease in systolic blood pressure was detected after administration of extract from Picrasma quassiodes, sesamin or silk fibroin hydrolysate, which improved antioxidative status [7,8,27]. However, treatment with the calcium antagonist nifedipine for 4 weeks did not induce any significant alterations in systolic blood pressure in stroke-prone spontaneously hypertensive rats (SHRSP), although it significantly reduced oxidative stress and inhibited intramyocardial arterial remodeling [10]. In the present study, the systolic blood pressure of SHR was maintained at a constant level after EEtRLP treatment for 2 weeks (Figure 2). These results are similar to the treatment effects of nifedipine. We believe that EEtRLP may contain a compound with nifedipine like function; however, further studies are needed to identify their structure and function.
eNOS in endothelial tissue participates in regulation of systolic blood pressure through generation of NO, smooth muscle contraction and plate aggregation [7,28,29]. In SHR, arterial dysfunction induced by the enhanced production of ROS causes NO breakdown and the suppression of eNOS expression [7]. However, the expression of eNOS is significantly recovered after treatment with several antioxidants. Treatment with sesamin (40-160 µg/kg) for 16 weeks enhanced the expression of eNOS in thoracic aortas [7]. Moreover, the number of positive particles of eNOS in the cytoplasm of vascular endothelial cells and smooth muscle cells was lower in the aorta of PQB-treated SHR than the vehicle-treated group. As shown in Figure 5, the expression of eNOS was dramatically increased by 4.2-8.3 times in the EEtRLP treated group. This pattern of eNOS expression detected after EEtRLP treatment was very similar to the results of previous studies; however, our findings constitute the first evidence that EEtRLP treatment can enhance eNOS expression in SHR aortas.
SOD has been reported to play an important role in the treatment of hypertension pathogenesis through elimination of oxidative stress. When SHR were in a hypertensive pathological state, SOD activity was significantly lower in the aorta compared to WKY rats [30]. However, a large number of animal studies have shown that antioxidant extract or compounds could induce enhancement of SOD activity in the hypertension model. Treatment with PQB at different doses improved SOD activity in the serum of SHR, while nifedipine selectively restored Cu/Zn SOD expression or activity in the WKY rat heart [10,23]. Moreover, SOD activity was gradually increased in SHR treated with increasing concentrations of silk fibroin hydrolysate [9]. In the present study, the activity of SOD in EEtRLP treated SHR was 300% greater than that of vehicle treated SHR. The increase in SOD activity in the EEtRLP treated group was about 46-168% higher than those of antioxidants investigated in previous studies.
Taken together, EEtRLP with high radical scavenging activity induced increased ACE expression and SOD activity while restoring the levels of aldosterone, K, MDA and NOx, although these compounds did not contribute to suppression of the systolic blood pressure. These results indicate that EEtRLP can relieve or prevent endothelial dysfunction, thereby reducing the incidence of hypertension and vascular disease.
Go to :
 XML Download
XML Download