

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Over the past decade, lutein has been recognized as an important dietary antioxidant for eye health, particularly for preventing age-related macular degeneration [1]. Among various carotenoids, lutein is a major plant oriented xanthophyll component that serves as a dietary antioxidant in the body [23]. Lutein's free radical scavenging antioxidant properties are known to play the principle role in preventing age-related macular disease since oxidative stress in retinal constituents is responsible for the pathogenesis of this disease [4]. Therefore, dietary lutein supplement is known to give rise to effective chain-breaking antioxidant activity due to its ability to quench singlet oxygen and scavenge peroxy radicals [56]. However, there is a still lack of evidence that dietary lutein can directly affect the induction of genes related to the antioxidant defense system in rats, although the major biological mechanism of lutein is linked to antioxidant activity.
Dietary supplementation of lutein is the only way to obtain lutein to prevent age-related cataracts and maculopathy, which are the major causes of blindness, since lutein is not synthesized within the body [7]. However, several studies have shown that the bioavailability and effectiveness of lutein varies greatly according to the complexity of the food matrix, interaction with other nutrients, ssolubility in the gut and dietary sources of lutein [89]. Several studies have reported that the lipid matrix of lutein showed better lutein absorption in the small intestine as a result of enhanced secretion of emulsifying bile acids by the gallbladder [91011]. Therefore, the beneficial effects of lutein are directly associated with the methods in which it is absorbed, deposited and metabolized in the body.
Several studies suggest that supplying mixtures of antioxidants including the carotenoids vitamin C or E may have more powerful antioxidant capacity than providing individual carotenoids [1213]. The mixture of β-carotene with vitamin C and E increased the activity of catalase and superoxide dismutase in humans remarkably [13]. The dietary mixture of carotenoids with vitamin C or folate showed remarkable benefits in cortical and posterior subcapsular cataracts in humans [14]. In contrast, lutein bioavailability was not affected by physiological concentration of vitamin C or E, but was partially hindered by the presence of β-carotene [15]. Therefore, definitive conclusions regarding the effects of dietary mixtures of antioxidants sources on the antioxidant defense system have not been reached, although the combination of several antioxidants appeared to have a synergistic effect on biomarkers of oxidative stress in humans and rats.
The primary goal of this study was to determine whether dietary lutein alone or in combination with vitamin C would influence the mRNA expression and activity of antioxidant enzymes, antioxidant capacity and lipid peroxidation in SD rats.
Materials and Methods
Experimental animals and procedures
The animal handling procedures were approved by the Institutional Animal Care and Use Committee (IACUC, No. 2014-2) at the Gyeongnam National University of Science and Technology in Korea. A total of thirty male Sprague Dawley (SD) rats purchased from Samtako Korea (Osan, Korea) were kept in a semi-barrier system with a regulated temperature (23±2℃) and light/dark cycle (light on 07:00-19:00). Immediately after a one week acclimation period, 18 rats with similar body weight were randomly allotted into three groups on the basis of body weight. All designated rats were housed in polycarbonate cages (two rats per cage) according to the experimental design. A commercial pelleted diet containing 23% crude protein, 5% crude fat, 5% crude fiber, and 45% nitrogen-free extract and sterilized bottle water were then provided ab libitum.
Each group was provided the pelleted diet and orally administered saline (CON), a commercial lutein dissolved into safflower oils at 50 mg lutein/kg diet (LUT), or a commercial lutein dissolved into safflower oils and vitamin C dissolved into water (50 mg lutein/kg BW and 0.5% vitamin C dissolved into H2O, LVC) for 4 weeks. Lutein and vitamin C were obtained from FloraGLO® (Lutein 20% SAF, DSM Nutritional Products Inc., Basel, Switzerland and Yuhan Pharmacy Co. (Products of DSM, UK)), respectively. A commercial lutein 20% supplement was supplied in an oil form on a 79% safflower oil carrier that contained 1% dl-α-tocopherol. A single dose of lutein was administrated orally daily using an oral zonde needle, while vitamin C was provided in a bottle of water containing 0.5% vitamin C (DSM, UK). The average daily intake of vitamin C was about 1,000 mg/kg BW per rat in the LVC group based on the water intake during the experimental period (data not shown).
Tissue harvesting
All rats were deprived of diets for 12 hours after the last day of the 4 week experimental period before administrating lutein. And then the rats (n=6 per group) were sacrificed with CO2 gas at 6 hours after gavage. Immediately after taking blood from the heart, the liver was harvested. The plasma and liver tissues were then rapidly frozen in liquid nitrogen and stored at -70℃ until further assay.
Plasma biochemical analyses
Plasma glucose, triglyceride, albumin, aspartate aminotransferase (AST) and alanine aminotranferease (ALT) were measured using a clinical biochemical analyzer (H1 System, Technicon, USA). All analyses were conducted in duplicate.
Expression of mRNA of antioxidant enzymes by Real Time-PCR
To analyze the mRNA expression of antioxidant enzymes including superoxide dismutase (SOD), glutathione peroxidase (GPX), glutathione S-transferase (GST) and catalase (CAT), mRNA was quantified by real-time PCR. The cDNA primers used to amplify each gene in this study are shown in Table 1. Total RNA from hepatic tissue was extracted using RNAzolTM B (TRIzol Reagent, Invitron, Eugene, OR, USA). Briefly, 2 mL of RNAzol solution was added to 100 mg of hepatic tissues from each individual. The tissues were then homogenized using liquid nitrogen and transferred to an Eppendorftube. Next, 100 µL of chloroform was added and incubated for 5 min on ice. The aqueous phase was then separated by centrifugation for 15 min at 15,000×g, after which the isolated RNA was precipitated with the same volume of isopropanol and then centrifuged for 15 min at 15,000×g. The separated total RNA was subsequently washed with 75% ethyl alcohol, dried, and diluted with diethylpyrocarbonate (DEPC)-treated H2O, after which the optical density at 260 nm was used to quantify the RNA (GeneQuant pro spectrophotometer, Amersham, Piscataway, NJ. USA). Next, 5 µg of total RNA were incubated with 1.0 µg of oligo dT (Invitrogen Co., Carlsbad, CA, USA) at 70℃ for 5 min and 4℃ for 5 min to synthesize the first strand cDNA. The reaction solution was incubated at 42℃ for 50 min, 90℃ for 10 min and 42℃ for 50 min in a reaction mixture containing 5× first strand buffer, 2.5 mM dNTP, 0.1M DTT, superscript III and RT-mixture. Quantification of the target genes and internal control (GADPH) was conducted by real-time quantitative PCR with SYBR green supermix (BioRad, Hercules, CA, USA) under the following conditions: 5 min at 95℃, followed by 40 cycles of denaturation at 95℃ for 15 s and annealing at 60℃ for 30 sec and then extension at 72℃ for 30 sec. The PCR amplification cycle at which dye fluorescence passed the selected baseline (Ct) was determined by realtime monitoring. The expression of all mRNAs was normalized by the 2[ΔΔ] method [16] to calculate relative changes in gene expression.
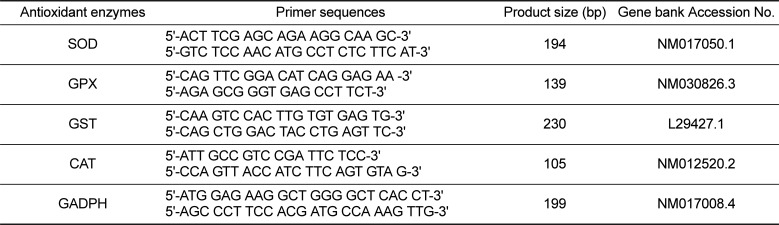
Table 1
Primers used for quantification of mRNA using Real Time-PCR
![]()
Assays of antioxidant enzyme activity, lipid peroxidation and total antioxidant capacity
To harvest the fraction of cytosol and microsomes, homogenized hepatic tissues were centrifuged at 10,000×g, after which the resulting supernatant was centrifuged at 105,000×g in a Centrikon T-2080 ultracentrifuge. The pellet (microsomes) was suspended in phosphate buffer (pH 7.4) to maintain a protein concentration of 20 mg/mL, after which the supernatant (cytosol) and a suspended pellet (microsomes) were frozen in liquid nitrogen and stored at -70℃ until further assay. The activity of SOD was analyzed in the cytosol using commercially available assay SOD assay kits (Sigma-Aldrich, St. Louis, MO, USA) based on an indirect assay method of xanthine oxidase as previously described [17]. The SOD activity was expressed as units/mg of proteins, where 1 unit of activity was the amount of enzyme required to inhibit 50% of the SOD or SOD like materials. GPX was measured at 37℃ in the cytosol with cumene hydroperoxide as a substrate [18]. The GPX coupled the reduction of cumene hydroperoxide to the oxidation of NADPH by glutathione reductase, and the concomitant oxidation rate was measured with a spectrophotometer based on the decrease in absorbance at 340 nm. One unit of GPX was expressed as the amount of GPX required to oxidize 1 µmol of NADPH per minute. GST was determined with 1-chloro-2,4-dinitrobenzene (CDNB) as a substrate by measuring the increase in absorbance at 340 nm [19]. One unit of activity was expressed as the amount of GST catalyzing the conjugated CDNB per minute. The lipid peroxidation level in the microsomes was determined by the concentration of thiobarbituric acid (TBA) reacting substances with a wavelength of 532 nm using a UV spectrophotometer [20]. TBA substances are described in nm of malondialdehyde (MDA) per mg of protein. Total antioxidant capacity in plasma and the cytosolic fraction of the liver was analyzed using a commercially available assay kit in conjunction with an ELISA reader (Oxford Biomedical Research, Inc. MI, USA). All assay procedures conducted exactly according to the manufacturer's protocols. Trolox was used to generate a standard curve and data were expressed as mM Trolox equivalents or in µM copper reducing equivalents. Protein concentration was assessed by the BCA method (Pierce Assay) using ELISA (VMax, Molecular Devices, CA, USA).
Statistical analysis
The effects of treatment on all data were analyzed by Proc GLM (SAS Institute Inc.). When the treatment effect was significant at P<0.05, Tukey's test was applied to assess significant differences among dietary groups. All values are presented as the mean±standard deviation (SD) and a P value of <0.05 was considered significant.
Go to : 
Results
Body and relative liver weights and blood biochemical parameters
Effects of the administration of saline (CON), lutein (LUT) or lutein in combination with vitamin C (LVC) on the body and the relative liver weights and blood biochemical parameters of SD rats are shown in Table 2 and 3, respectively. After 28 days of treatment, no significant difference in body and relative liver weights was observed among treatment groups (Table 2). In addition, the levels of blood parameters including glucose, triglyceride, albumin, AST and ALT in the LUT and LVC groups were very similar to those of the CON group (Table 3).

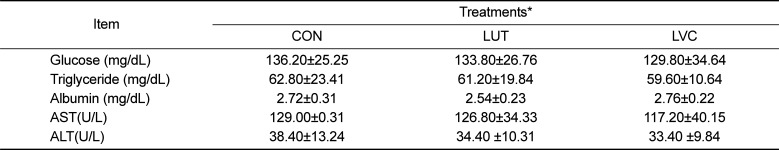
Table 2
Changes in body weight and relative live weight of SD rats administrated saline (CON), lutein (LUT) and lutein in combination with vitamin C (LVC)
![]()
Table 3
Changes in blood biochemical profiles of SD rats administrated saline (CON), lutein (LUT) and lutein in combination with vitamin C (LVC)
![]()
mRNA expression and activity of antioxidant enzymes in the liver
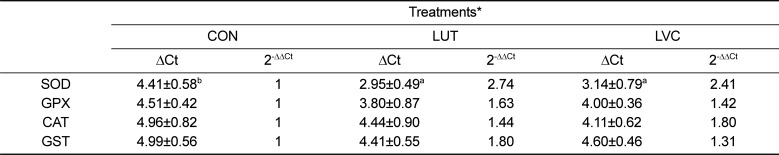
The mRNA expression level and activity of hepatic SOD, GST, GPX and CAT in the CON, LUT and LVC groups is presented in Table 4 and Figure 1, respectively. The LVC group showed a significantly (P<0.05) greater mRNA expression of SOD than the CON group. However, mRNA expression of GPX, GST and CAT was not affected by the administration of lutein or lutein plus vitamin C in the liver of SD rats (Table 4). Similar to the pattern of mRNA expression, the activity of hepatic SOD in the LUC and LVC groups was significantly (P<0.05) higher than that of the CON group, whereas the GPX and CAT activities were not influenced by the administration of lutein or lutein in combination with vitamin C (Figure 1). The activity of GST was significantly (P<0.05) higher in rats administrated lutein (LUT) or lutein in combination with vitamin C (LVC) than in control rats (CON).
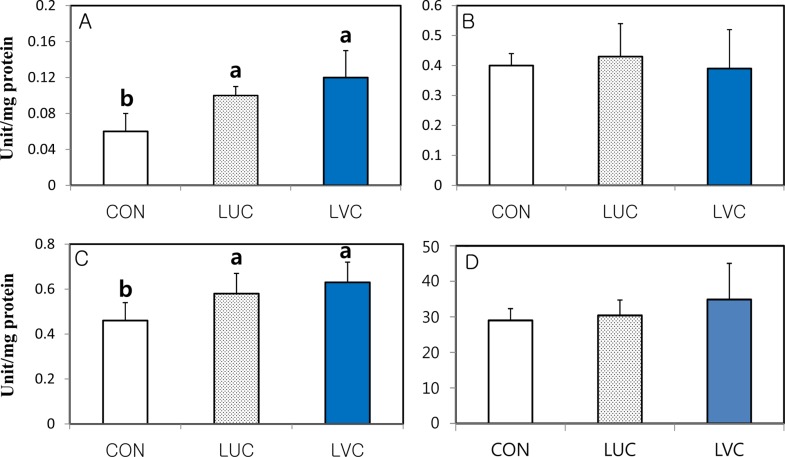 | Figure 1Specific activity of antioxidant enzymes (A: SOD, B: GPX and C: GST and D: CAT) in the liver of SD rats administrated saline (CON), lutein (LUT) and lutein in combination with vitamin C (LVC). Means (Mean±SD, n=5) with different superscript differ among dietary groups (P<0.05).
|
Table 4
Changes in mRNA expression of antioxidant enzymes in the liver of SD rats administrated saline (CON), lutein (LUT) and lutein in combination with vitamin C (LVC)
*CON (saline), LUT (50 mg lutein/kg BW) and LVC (50 mg lutein and 1,000 mg vitamin C/kg, BW).
The values are ΔCt, which is represneted as the Ct of each target gene corrected by Ct of the control gene (GADPH).
The fold difference in the relative expression of the target gene was calculated as the 2-ΔΔCt.
Means (Mean±SD, n=5) with different superscript differ among dietary groups (P<0.05).
![]()
Total antioxidant power and lipid peroxidation in the blood and liver
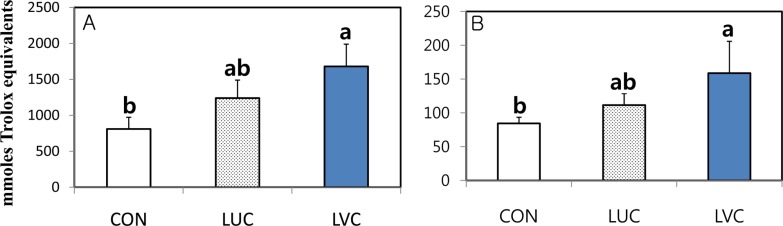
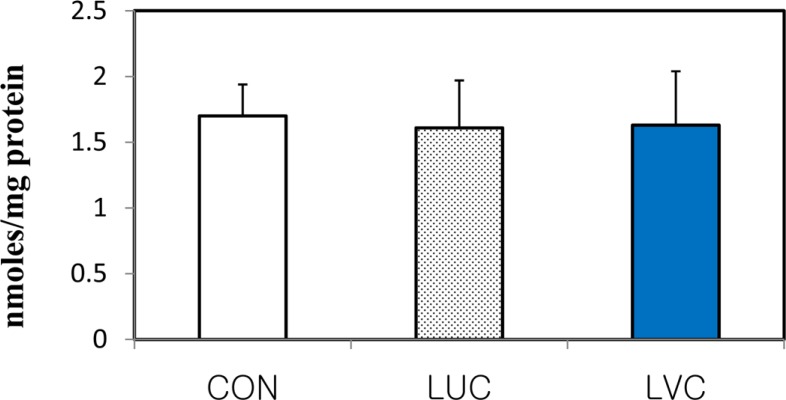
To determine whether a dietary source of antioxidant affects the antioxidant defense system in the plasma and liver of rats, we examined the level of total antioxidant power (TAP) and lipid peroxidation (Figures 2, 3). The total antioxidant power in the plasma and liver significantly (P<0.05) increased in groups administrated lutein in combination with vitamin C relative to the control group (Figure 2). However, lipid peroxidation as indicated by MDA level in liver tissues of rats was unaffected by the administration of lutein or lutein in combination with vitamin C (Figure 3). Although lipid peroxidation of plasma and liver tissues did not differ significantly among groups, the LVC group showed a significantly higher level of total antioxidant capacity compared with the CON group, indicating that the administration of lutein in combination with vitamin C could effectively protect against reactive oxygen radicals in rats.
Go to :
Discussion
In this study, we investigated the body and liver weights and blood biochemical profiles of SD rats after administration of lutein or lutein in combination with vitamin C. The results showed that treatment with lutein or lutein with vitamin C did not induce changes in glucose, triglyceride, albumin, AST and ALT levels in SD rats. Measurements of AST and ALT have been reported as indirect methods to assess liver damage [21]. Similar to the present study, a study reported that plasma AST and ALT did not differ between the control and lutein groups, although lutein has been shown to decrease oxidative stress and inflammation in the liver and eyes of guinea pigs fed a hypercholesterolemic diet [22].
To evaluate the effects of lutein alone or in combination with vitamin C on the antioxidant defense system in rats, we examined the mRNA expression and activity of antioxidant enzymes, lipid peroxidation and total antioxidant capacity. Rats that received lutein in combination with vitamin C showed the highest level of antioxidant activity, including increased mRNA expression and activity of SOD and total antioxidant capacity in plasma and liver tissues. Therefore, it is thought that increased SOD and total antioxidant capacity in response to supplying lutein in combination with vitamin C might act as powerful antioxidant scavengers against oxidative stress in the tissues of rats. These findings also suggest that vitamin C has a synergistic effect on the antioxidant defense system in rats administered lutein. The administration of lutein alone at the level of 50 mg/kg BW was not sufficient to increase total antioxidant capacity in the plasma and liver, although rats that received lutein alone showed increased induction of some antioxidant enzymes.
Carotenoids such as lutein, a family of fat-soluble pigments, are known to be effective chain-breaking antioxidants due to their ability to scavenge reactive oxygen radicals [6]. However, there is still a lack of evidence that dietary lutein can modulate endogenous antioxidant enzymes in rats, although numerous studies have reported that lutein showed beneficial effects on antioxidant capacity against reactive oxygen species (ROS) in vitro [2324].
In partial agreement with the results of the present study, mice orally administered lutein (50, 100 and 200 mg lutein/kg of BW) showed significantly increased activities of SOD, GST and GPX in the liver [25]. The oral administration of lutein (0.2 and 0.5 mg lutein/kg of BW) in mice significantly enhanced the mRNA expression of SOD and GPX in the liver without affecting lipid peroxidation [26], suggesting that lutein might induce the expression of antioxidant genes. In contrast, several studies indicated that dietary lutein did not directly affect the induction of antioxidant enzymes, including SOD and GPX, in humans and rats [2728]. In addition, dietary supplementation of lutein to rats (20 mg/kg, BW) had no effect on the expression and activity of serum SOD, GPX and CAT compared to control rats, whereas lutein supplementation reversed the noticeably-decreased antioxidant enzyme activity of rats exposed to severe oxidative stress [29]. Interestingly, dietary lutein attenuates hepatic oxidative damage via activation of antioxidant enzymes under severe oxidative stress conditions [1129], suggesting that dietary lutein has antioxidant ability under severe oxidative stress. In agreement with this observation, administration of lutein to rats activated antioxidant enzymes including SOD, GPX or CAT when cellular damage was induced by oxidants [30]. A study also demonstrated that antioxidant enzymes such as SOD and GPX were enhanced in lutein-fed rats when compared with control rats when hepatic damage was induced by alcohol and CCL4 [31]. Moreover, supplementation of the diets of rats with lutein under severe oxidative stress significantly improved the antioxidant defense system by directly scavenging ROS and indirectly activating antioxidant enzymes [26].
Antioxidant vitamins including Vitamin C or E are well known to exert antioxidant capacity against ROS induced by oxidative stress in humans and rodents [3233]. Vitamin C as an electron donor is a well-known redox catalyst that can reduce and thereby neutralize ROS such as hydrogen peroxide [14]. Vitamin E is a very important lipid-soluble antioxidant that is also effective at chain-breaking, enabling protection of membranes from oxidation by reacting with lipid radicals generated during lipid peroxidation [3334]. Recently, evidence has emerged showing that these vitamins interacted with carotenoids to modulate antioxidant status. The mixture of lutein with vitamin C or E showed a significantly positive synergistic effect on the biomarkers of oxidative stress in rats [1214]. In this study, the mRNA expression and activity of SOD in the liver was more significantly enhanced when rats were given lutein in combination with vitamin C, indicating that the mixture of lutein with vitamin C has a potentiating effect relative to single carotenoids. In accordance with our study, the antioxidant effects were more potent when lutein was supplied with vitamin C rather than with vitamin E in Zucker rats [14]. The combined effects of lutein with vitamin C on biomarkers of oxidative stress were also shown to be more potent than those of lutein administered with vitamin E [14]. Dietary supplementation of both β-carotene and vitamin C significantly enhanced the activity of catalase and SOD in humans [13]. A recent study demonstrated that there was a significant association between plasma lutein and vitamin C levels and longer telomere length in humans, suggesting that lutein and vitamin C play a significant role in protecting the integrity of the telomere from oxidative stress [35]. Since dietary total antioxidant capacity is directly related to plasma antioxidant status in humans [36], evidence from our previous studies indicates that lutein interacts with antioxidant vitamins such as vitamin C to maintain the antioxidant defense system in rats.
One possible reason for the synergistic effects of lutein administered in combination with vitamin C is that lutein absorption may be activated by the presence of vitamin C. Interestingly, ddietary vitamin C supplementation significantly facilitated dietary lutein absorption in humans [37]. However, few studies have investigated whether dietary supplementation with vitamin C could have potentiated the absorption of lutein, even though the combination of lutein and vitamin C appears to have potentiating effects on biomarkers of oxidative stress in humans and rats.
Overall, the dietary supplementation of lutein in combination with vitamin C showed more consistent effects and greater significance in the antioxidant defense system than that of lutein alone, although lutein only has been reported to have potential antioxidant capacity. Therefore, our results indicate that lutein in combination with vitamin C may have more beneficial effects on oxidative stability through modulation of the expression of antioxidant enzymes and total antioxidant capacity than lutein alone. However, more sophisticated studies are still needed to elucidate the antioxidant mechanism that enables lutein in combination with vitamin C to cope with oxidative stress conditions.
Go to :
 XML Download
XML Download