

 PDF
PDF ePub
ePub Citation
Citation Print
Print


According to the change in the pattern of food consumption, the incidence of metabolic syndrome such as obesity, atherosclerosis, hypertension, stroke, diabetes, and cancer are considerably increasing [1,2]. Especially, hyperlipidemia might be the main cause of health problems in metabolic syndrome [3].
The major risk factors of cardiovascular diseases are increasing levels of total cholesterol and low-density lipoproteins (LDL), accompanying decrease in high-density lipoproteins (HDL) [4,5]. LDL as a major transporter of blood cholesterol is oxidized in the endothelium of arterial walls and induces infiltration of circling monocytes following endothelial cell injuries. In contrast, HDL transports cholesterol to the liver and facilitates its binding to LDL receptors for metabolism [6,7,8,9], Oxidized LDL (OxLDL) stimulates vascular endothelial cells to produce monocyte-colony stimulating factor (M-CSF) and induces differentiation of infiltrated monocytes to macrophages [8]. Then, LDL binds to several scavenger receptors expressed on macrophages instead of LDL receptors, disrupting feedback controlling mechanism of intracellular cholesterol level. Macrophages that contain highly enriched cholesterol become foam cells and form atheromatous plaques, initial lesion of atherosclerosis [9].
Vascular smooth muscle cell (VSMC) proliferation and neointimal formation are major events in the pathophysiology of atherosclerosis. After vascular injury, resting VSMCs, in a non-proliferative quiescent state, loose a well-differentiated contractile phenotype, shifting to a synthetic phenotype, entering the cell cycle and proliferation [10,11]. Platelet-derived growth factor (PDGF)-BB, which is a classic growth factor secreted by injured endothelial cells and VSMCs as well as by platelets and macrophages, promotes the proliferation of fibroblasts, glia, and VSMCs [12]. PDGF-BB propagates mitogenic signals through autophosphorylation of its respective PDGF β-receptor on tyrosine residues, thus triggers downstream signal transductions and cell cycle progression [13,14,15]. Proliferation of VSMCs plays a key role for thickening of arterial walls, narrowing the lumen of vessels and increasing tension.
Statins inhibit 3-hydroxy-3-methylglutaryl-coenzyme A (HMG-CoA) reductase, rate-limiting step enzyme in cholesterol biosynthesis, thereby reduce intracellular cholesterol level. Statins also increase the activity of LDL receptors on cell membrane, accelerating the metabolism of very low-density lipoproteins (VLDL) remnant and LDL [16,17]. Even though drugs with such mechanism have an advantage to lower blood cholesterol level remarkably, but they exert serious adverse effects like hepatomegaly and renal hypertrophy [18,19]. Thus, researches on the development of natural resources with a novel mechanism without side effects are required.
Traditionally, the dried roots of Angelica gigas Nakai (Korean angelica), known by the Korean name as 'Cham-dang-gui', have been used in Oriental medicine for the treatment of anemia and some circulatory disorders [20]. Several coumarin derivatives, such as decursins, decursinol, decursinol angelate, nodakenin, nodakenetin, and umbelliferone, were isolated from Angelica gigas [21,22,23]. Additional pharmacological effects of these derivatives have been reported: decursinol from Angelica gigas was shown to possess anti-nociceptive activity [24], and decursin inhibited the induction of inflammatory mediators from lipopoly-saccharide-stimulated macrophages [25]. Recently, we demonstrated that an ethanolic extract of Angelica gigas (EAG) has anti-inflammatory activities in carrageenan-induced air-pouch inflammation, croton oil-induced ear inflammation, and dinitrofluorobenzene-induced allergic dermatitis models [26,27,28]. In addition, Lee et al. [29] showed the inhibitory activity of coumarin derivatives from Angelica on the platelet aggregation as a rationale for the traditional use in anemia and some circulatory disorders [20].
Therefore, such results led us to investigate the effects of EAG on the PDGF-BB-stimulated VSMC proliferation and high-cholesterol diet (HCD)-induced hypercholesterolemia and atherosclerosis.
The root of Angelica gigas was purchased from a local Oriental medicine market in Seoul, Korea in 2012. The plant materials were authenticated by an expert at Chungbuk National University. The dried root (5 kg) of Angelica gigas was extracted with 70% ethanol or distilled water in an ultrasonic apparatus [26,27,28]. Upon removal of the solvent under vacuum, the ethanolic extract (EAG) and water extract (WAG) of Angelica gigas yielded 500 g and 670 g, respectively.
Seven-week-old male Sprague-Dawley rats (250-300 g) and six-month-old male New Zealand white (NZW) rabbits (1.8-2.0 kg) were procured from Daehan-Biolink (Eumseong, Korea), and subjected to the experiment after 1-week acclimation to the laboratory environment. The animals were housed in each cage with free access to feed and water under constant environmental conditions (22±2℃; 40-70% relative humidity; 12-hour light-dark cycle; 150-300 lux brightness). All the animal experiments were conducted according to the Standard Operation Procedures, and approved by the Institutional Animal Care and Use Committee of Chungbuk National University, Korea.
Rat aortic VSMCs were isolated by enzymatic dispersion as previously described [30]. Cells were cultured in Dulbecco's Modified Eagle Medium (DMEM), supplemented with 10% fetal bovine serum (FBS) [31,32]. The purity of VSMCs culture was confirmed by immunocytochemical localization of α-smooth-muscle actin.
The proliferation of rat aortic VSMCs was measured by direct counting. The VSMCs were seeded into 12-well culture plates at 4×104 cells/mL, and then cultured in DMEM containing 10% FBS at 37℃ for 24 hours. After reaching at ~70% of confluence, the cells were incubated with serum-free medium for 24 hours, treated with various concentrations (1-10 µg/mL) of EAG for another 24 hours in newly fresh serum-free medium and stimulated by PDGF-BB (25 ng/mL) [31,32]. After 24 hours, the cells were trypsinized by trypsin-EDTA and counted by using a hemacytometer under a microscope.
DNA synthesis in VSMCs was determined by [3H]-thymidine incorporation assay as previously described [31,33]. The assay condition was the same as described in the section of cell proliferation assay. Under the stimulatory condition of PDGF-BB (25 ng/mL) addition in serum-free medium, [3H]-thymidine (2 µCi/mL) was added for 4 hours before harvesting. The reaction was terminated by aspirating the medium and subjecting the cultures to sequential washes on ice with PBS containing 10% trichloroacetic acid and ethanol/ether (1:1, v/v). Acid-insoluble [3H]-thymidine was extracted into 250 mL of 0.5 M NaOH/well, and this solution was mixed with 3 mL scintillation cocktail (Ultimagold, Packard Bioscience, CT, USA), and quantified using a liquid scintillation counter (LS3801, Beckman, Düsseldorf, Germany).
Acute dietary hypercholesterolemia was induced by feeding rabbits with a powdered high-cholesterol diet (HCD) containing 0.5% cholesterol for 2 weeks, followed by 0.5% cholesterol for additional 8 weeks during treatment period [34,35]. After 2-week induction period, the animals were grouped (n=6/group) according to their blood cholesterol levels to adjust similar mean values. To assess therapeutic efficacy of EAG against hypercholesterolemia and atherosclerosis, the rabbits were fed the HCD containing 1% EAG for 8 weeks.
After 16-hour fasting at the end of 2-week hypercholesterolemia induction and every 2 weeks during 8-week experimental period, blood sample was collected from auricular artery, and lipid profiles including total cholesterol, LDL, and HDL were measured in sera using a blood chemistry analyzer (Hitachi-747; Hitachi Korea, Seoul, Korea).
The aortic arch (10 cm) was dissected from the aortic valve of the heart and washed with saline. The sample was dissected in 5 cm long from the orifice of carotid artery, and opened longitudinally. After removing fats and tissues adhering to the adventitia, the aorta was dehydrated with 100% propylene glycol for 10 min, and stained with 0.7% Sudan IV (in propylene glycol) for 10 min [34,35]. Then, it was hydrated with 85% propylene glycol for 5 min, washed with distilled water, and photographed and analyzed with Digital Image Analyzer (Image Inside; Focus, Seoul, Korea) for red atheromatous plaques. The extent of lipid accumulation (atherosclerosis index; AI, %) was calculated as the percent Sudan-positive area to the total area of the aortic wall.
The results were presented as mean±standard error. The significance of differences of all results was analyzed by one-way analysis of variance followed by the Dunnett's multiple-range test correction. Statistical significance was set a priority at P<0.05.
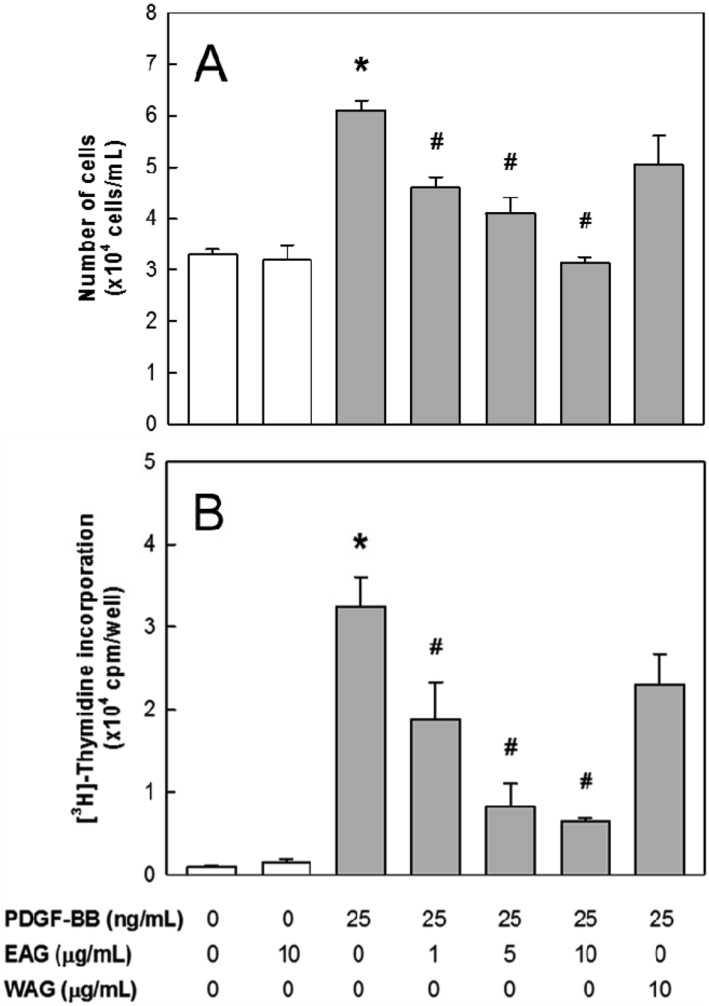
PDGF-BB (25 ng/mL) induced VSMC proliferation, doubling the number of cells in 24 hours (Figure 1A). EAG significantly inhibited the PDGF-BB-induced cell proliferation in a concentration-dependent manner, decreasing the number by 50% and 100% at 1 and 10 µg/mL, respectively. In parallel with the suppressed proliferation, the thymidine incorporation in VSMCs was inhibited by EAG (Figure 1B). By comparison, the effects of WAG on the proliferation and DNA synthesis were much lower than EAG (Figure 1). Separately, EAG did not affect both the proliferation and thymidine incorporation of normal (PDGF-BB-unstimulated) VSMCs. According to the relatively-low activity on VSMC proliferation, WAG was excluded for further studies on antherosclerosis.
After 2-week feeding of 0.5% cholesterol, mean blood cholesterol was dramatically elevated to 1,081 mg/dL from 60 mg/dL of normal level (Figure 2A). Accordingly, LDL also increased to 313 mg/dL from 22 mg/dL in normal animals, although there was a relatively-low increase in HDL levels to 39 mg/dL from 30 mg/dL (Figures 2B and 2C). Feeding EAG slightly decreased both the cholesterol and LDL levels, but significantly increased (preserved) HDL level.
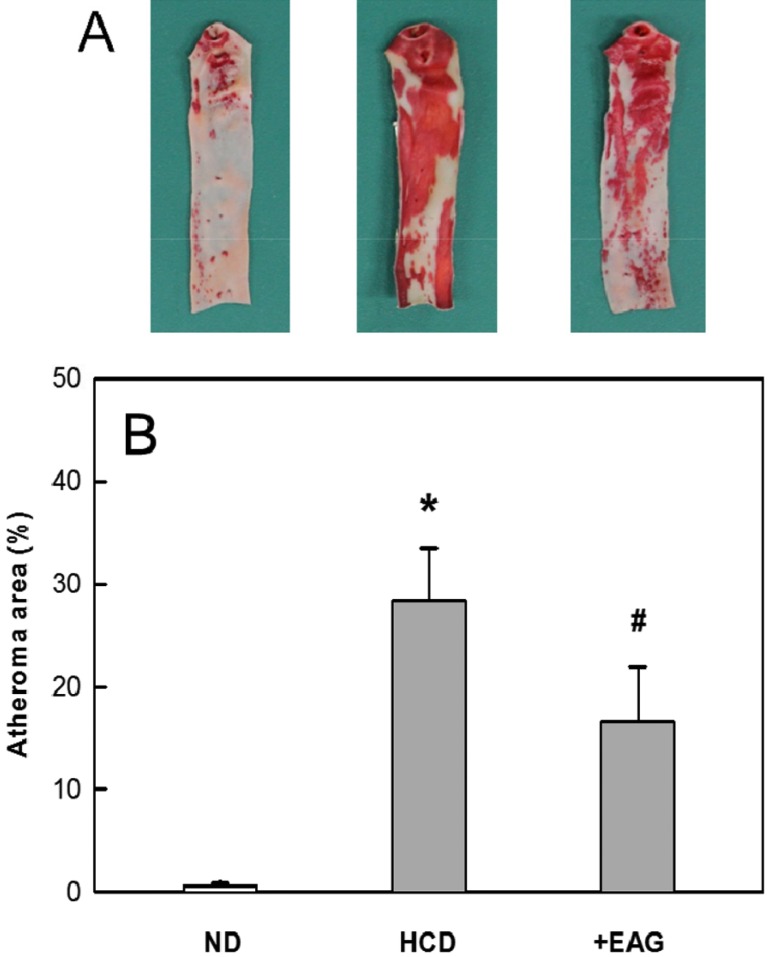
Ten-week feeding 0.5% cholesterol caused extensive atheromatous plaques covering 28.4% of the aortic arch and abdominal artery (Figure 3). The degree of atheromatous plaques formation was attenuated by 8-week treatment with EAG (1% in diet) to 16.6%.
VSMCs are the main component of arterial wall and in a non-proliferative quiescent state in normal artery, and show a well-differentiated contractile phenotype. After vascular injury, there is a loss of differentiated phenotype and a shift to a synthetic phenotype, which is accompanied by entry into the cell cycle and proliferation, and neointimal formation. The abnormal proliferation of VSMCs plays a central role in the pathogenesis of atherosclerosis and restenosis after balloon angioplasty, and possibly in the development of hypertension [10,11,36]. Interestingly, our results described that the treatment of EAG on rat aortic VSMCs drastically decreased their proliferation stimulated by PDGF-BB, fully suppressing at a low concentration (10 µg/mL). Moreover, such an anti-proliferative activity of EAG was confirmed due to EAG's inhibitory potential on the DNA synthesis. Therefore, it is inferred that EAG strongly decreases neointima formation. Since the accumulation and proliferation of VSMCs within the intimal layer of vessel wall is known to be a key step in the development of atherosclerotic plaques, EAG may have a preventive role in atherosclerosis [37]. Since EAG did not affect the proliferation and thymidine incorporation of unstimulated VSMCs, the anti-proliferative effect was not due to cellular cytotoxicity or apoptosis.
Based on the beneficial effects of EAG on blood flow, i.e., anti-proliferation of VSMCs (the present study), anti-platelet aggregation [29], and anti-inflammation [25,26,27,28], it was expected that long-term feeding of EAG should inhibit atherosclerosis. Actually, EAG (1% in diet) significantly inhibited the atheroma formation in the aortic walls of HCD-fed rabbits. It is of interest to note that the improving activity of EAG on the lipid profiles of the hypercholesterolemic rabbits was not enough to explain the marked anti-atherosclerotic effects. Therefore, it is assumed that the anti-atheromatous activity of EAG might be from its potential for the inhibition of VSMC proliferation.
Although HMG-CoA reductase inhibitors such as lovastatin is preferentially prescribed for hypercholesterolemia and cardiovascular diseases, because they suppress cholesterol synthesis and improve bile duct secretion, remarkably improving blood lipid profiles [38,39], it is well known that long-term administration of the statins cause severe hepatotoxicity [18,35]. Many natural products such as spirulina possessing anti-cholesterolemic potential without serious adverse effects have been proposed to be good alternatives for the highly-toxic drugs [34]. Notably, EAG exerted anti-atherosclerotic effects by inhibiting VSMC proliferation, which is a novel action mechanism different from other natural products. Therefore, it is suggested that EAG could be an alternative or an adjunct therapy for the improvement of hypercholesterolemia and prevention of atherosclerosis.

 XML Download
XML Download