

 PDF
PDF ePub
ePub Citation
Citation Print
Print


The pancreas is a complex gland having both exocrine and endocrine activities. The exocrine pancreas in the mature animal synthesizes and stores digestive enzymes to release after ingestion of a meal that induces neural and hormonal stimulation [1]. The endocrine pancreas contains five major cell types, which produce insulin (B cells), glucagon (A cells), somatostatin (D cells), pancreatic polypeptide (PP cells) and ghrelin, which is a novel growth hormone-releasing peptide (ghrelin cells). B cells may also express islet amyloid polypeptide (IAPP), another member of the insulin family of proteins, insulin-like growth factor-II (IGF-II) and pituitary adenylate cyclase activating polypeptide (PACAP) [2].
The pancreas releases these various enzymes and hormones to digest foods and maintain glucose homeostasis. If pancreas is damaged, the aftermath can be serious. Two examples are diabetes mellitus and pancreatic adenocarcinoma. Diabetes mellitus is a chronic metabolic disorder resulting from defects in insulin secretion or function. The global prevalence of diabetes was estimated as 2.8% in 2000 and is predicted to grow to 4.4% by 2030. The total number of diabetics is projected to rise from 171 million in 2000 to 366 million in 2030 due to population growth, aging, urbanization and increasing prevalence of obesity and physical inactivity [3]. Pancreatic adenocarcinoma is the fourth leading cause of cancer deaths in the United States and its prognosis is usually dismal [1].
Preventive and effective treatments of pancreas-related diseases including diabetes mellitus, pancreatitis and pancreatic cancer require knowledge of the biology and molecular characteristics of the pancreas. Better understanding of pancreatic development and regeneration will provide valuable information about the identification of molecular markers involved in the pathogenesis of pancreatic cancer. As well, marker identification can be useful for islet cell transplantation, which is being explored as a treatment for diabetes.
Few proteomic studies have been done using human pancreatic tissues to elucidate the mechanisms of diabetes mellitus and pancreatic cancers because of difficulty in preparing human samples [4,5,6,7]. Instead, in vivo experiments have mainly used animal models. Several studies have contributed to the understanding of the molecular network governing developing pancreas in mouse and rat models [8,9].
The pig has become an attractive subject of study because of its potential use for xenotransplantation [10]. Nonhuman primate, such as apes and monkeys, more closely resemble humans anatomically and physiologically than do pigs. Nonetheless, xenotransplantation using nonhuman primates as xenograft donors is unlikely primarily because of the infectious risks to human patients and their contacts. Pigs produce large litters (up to 10 littermates), have a short gestation time (4 months), are somewhat similar to humans anatomically and physiologically, are available in large numbers (an estimated 90 million pigs are raised for human consumption yearly in the United States) and have a long history of providing medicinal products (skin, insulin, cardiac prostheses, clotting factors) for humans. For these reasons, the pig has become the most likely candidate for consideration as an organ donor [11,12,13,14]. Hindering this potential, the development of pig pancreas is not well understood.
In this study we investigated changes in the proteomic profile during pig pancreas development in embryonic day 60 (E60), neonatal and 6-month-old pigs using two-dimensional gel electrophoresis (2-DE) and matrix-assisted laser desorption/ionization-time of flight mass spectrometry (MALDI-TOF MS).
Materials and Methods
Pancreas protein extraction
Pancreas of E60 (n=4), neonate (n=3), 6-month-old male pigs (n=3) were removed immediately after sacrifice and stored in liquid nitrogen until used. Frozen pancreas tissues were homogenized in 400 µl lysis buffer (7 M urea, 2M thiourea, 2% w/v CHAPS, 2% Pharmalyte pH3-10, 100 mM DTE). After centrifugation (50,000 rpm at 4℃ for 1 h) the supernatant was extracted and complete protease inhibitor cocktail (Roche Applied Science, Basel,, Germany), DNase I (2.5 mg/mL; Roche Applied Science) and RNase (2.5mg/mL; Roche Applied Science) were added. The protein concentration was measured by spectrophotometry using Bio-Rad Protein Assay (Bio-Rad Laboratories, Hercules, CA, USA) based on the Bradford method. Because urea, thiourea, CHAPS and other constituents of the sample/lysis buffer can interfere with the protein estimation, the standard curve was generated using bovine serum albumin (BSA) at concentrations of 0, 2, 5, 7 and 10 mg/mL. In this process, pancreas sample was made from a pool of each of the 3~4 animals in each group, and each pooled sample was run 3 times.
2-DE
2-DE was performed as previously described [15]. One milligram quantities of protein were diluted in the sample buffer with 2% IPG buffer up to 450 µL. Samples were added to pH 3-10, nonlinear, 24 cm IPG strips (Amersham Pharmacia Biotech, Piscataway, NJ, USA) and covered with mineral oil. After rehydration for 16 h at 20℃, isoelectric focusing was carried out in eight steps under step-n-hold mode as follows: (S1) 30 V, 2 h; (S2) 100 V, 2 h; (S3) 200 V, 1 h; (S4) 500 V, 1 h; (S5) 1,000 V, 1 h; (S6) gradient, 8,000 V, 1 h; (S7) gradient, 100,000 V, 1 h and (S8) 20,000 VHrs. The total voltage applied was 129,960 Vh (IPGphore Isoelectric Focusing Unit: Amersham Pharmacia Biotech). The IPG strips were equilibrated in buffer containing 6M urea, 20% glycerol, 1.5 M Tris-HCl, pH 8.8, 2.5% acrylamide and Tributyl phosphine (Fluka, Buchs SG, Switzerland). After equilibration, SDS-PAGE was performed by 8-18% gradient gel without stacking gel via the Ettan Dalt system (Amersham Phamacia Biotech). 2-DE was carried out overnight at 4W per gel at 20℃. After finishing 2-DE, the proteins on gels were fixed with protein fixing solution (40% methanol, 5% phosphoric acid) for 2 h and then stained using Coomassie G-250 (Bio-Rad) for 3 h. To remove extra dye in gels, the gels were washed with distilled water.
Protein visualization and image analysis
The stained gels were scanned using a GS-800 calibrated densitometer (Bio-Rad) and spot analysis was performed with the ImageMaster™ 2D platinum software version 5.0 (Amersham Pharmacia Biotech). The digitalized 2-DE gel images were compared by matching using Image master 5.0 (Amersham Biosciences, Piscataway, NJ, USA).
In-gel digestion and protein identification by MALDI-TOF MS
Spots of interest on the 2-DE gel were excised, cut into small pieces and digested with 12.5 ng/µL trypsin (Promega, Madison, WI, USA) in 50 mM ammonium bicarbonate (pH 8.0), as previously described [16]. The preparation procedures for MALDI-TOF MS used reversed-phase columns as previously described [17]. The tryptic peptides were passed through a POROS 50 R2 column (Applied Biosystems, Forster City, CA, USA). After washing the column with 70% acetonitrile in 5% formic acid, 100% acetonitrile and 5% formic acid, peptides were loaded into column and washed with 5% formic acid. The peptides were eluted with 10 mg/mL a-cyano-4-hydroxy-cinnamic acid (Sigma-Aldrich, St. Louis, MO, USA) containing solution and elutes were dropped on a MALDI sample plate [18]. Mass analyses were performed using a Voyger-DE PRO MALDI-TOF MS apparatus (Applied Biosystems, Franklin Lakes, NJ, USA), equipped with a 337 nm nitrogen laser. This machine was operated at an accelerating voltage of 20 K, positive ion reflection mode, voltage grid of 74.5%, guide wire voltage of 0% and a delay time of 110 ns. The spectra were externally calibrated using a standard peptide mixture containing angiotensin I, 1,296.68 [M+H]; Glu-Hibrinopeptide B, 1,570.67 [M+H]; ACTH (clip18-39), 2,465.19[M+H]). The proteins were identified by Swiss-Prot and MSDB protein database using MS-FIT (Protein Prospector; UCSF, San Francisco, CA, USA). All the cases of search were analyzed with a 100 ppm mass tolerance and 1 miss cleavage.
Results
Protein identification and analysis during pancreas developmental stages
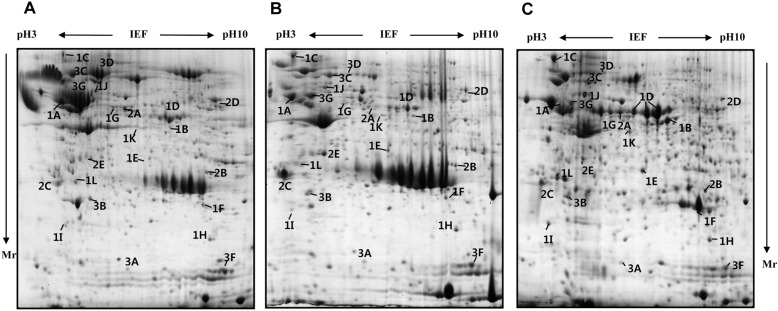
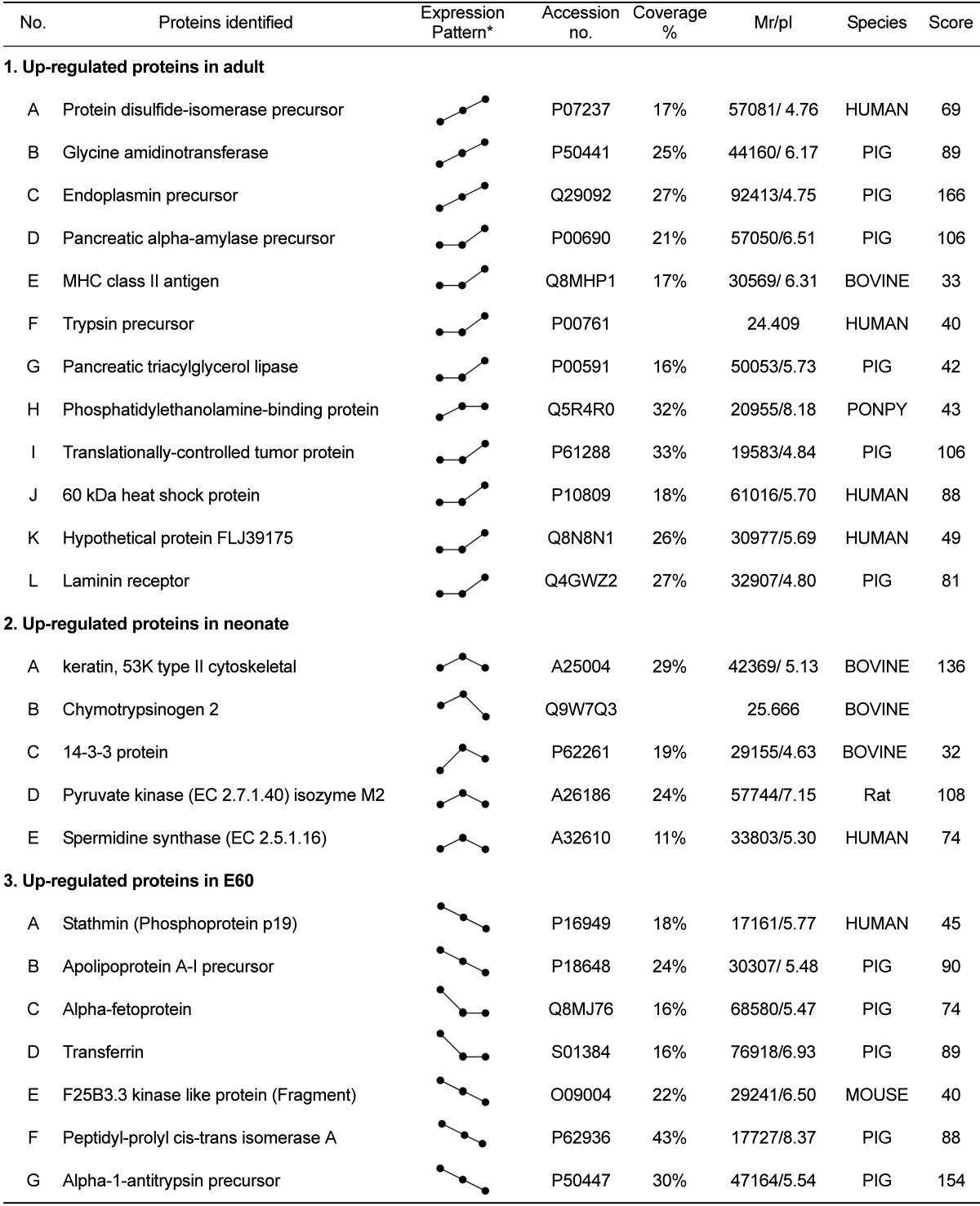
To analyze the changes in the proteome profile during pig pancreas development, the pancreas of E60, neonate, and 6 month-old pigs were used. Pancreatic tissue was processed for 2-D electrophoresis to isolate the candidate proteins that were differentially expressed during development. The change of protein expression was detected in each of the Coomassie blue stained gels using computer-assisted image analysis. At each respective developmental stage, a total of 467, 402 and 557 spots were detected from each gel (Figure 1). Among them, 24 proteins showed marked changes in expression. Twelve spots increased, while 7 spots decreased. The expression level of 5 spots was highest at birth. Twenty four differentially expressed proteins were identified by MALDI-TOF MS and MASCOT (Table 1). Changes in protein expression patterns were visualized in gels.
Up-regulated proteins in adult
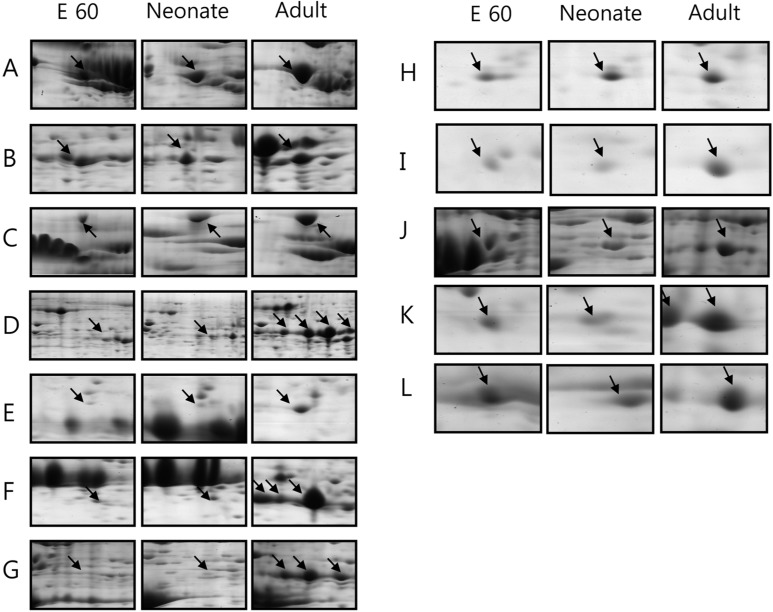
The amount of protein disulfide-isomerase precursor, endoplasmic precursor, glycine aminotransferase and heat shock protein increased gradually during the development from fetus to adult pig (Figure 2). Their increase suggests that gradual increase in body mass and food uptake of animal results in higher protein content of these proteins for making the necessary quantities of digestive enzymes and hormones. In addition, digestive enzyme precursors, such as pancreatic alpha-amylase precursor, trypsin precursor and pancreatic triacylglycerol lipase, were stage-specific proteins dominantly expressed in adult pig pancreas. Immune-mediated MHC class II antigen was expressed in large quantities in the adult pancreas.
Up-regulated neonatal proteins
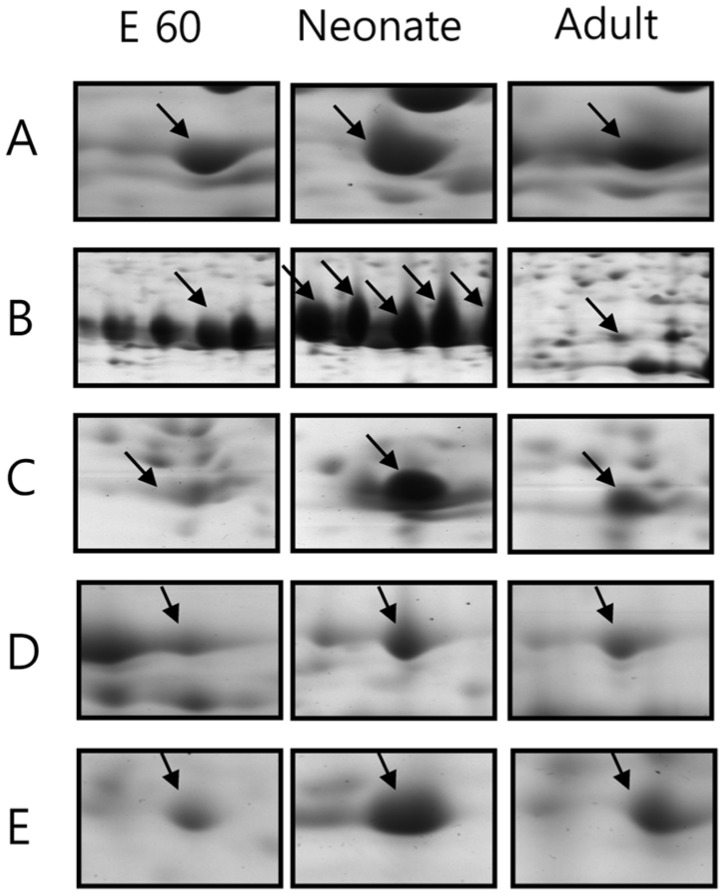
Expression of 5 proteins were up-regulated at the time of birth: chymotrypsinogen 2, pyruvate kinase isozyme M2, spermidine synthase and 14-3-3 protein (Figure 3). Among them, the change in expression pattern of chymotrypsinogen 2 was especially pronounced. However, although chymotrypsinogen 2 was rarely expressed in adult pig pancreas, it occupied at least 20% of entire pancreas proteins in fetus and neonate, especially neonates. The specific activity of chymotrypsin increased until birth and then declined precipitously, consistent with prior reports [19,20].The activities of other zymogens (amylase, lipase and elastase) were low in the fetus and steadily increased after birth.
Up-regulated fetal proteins
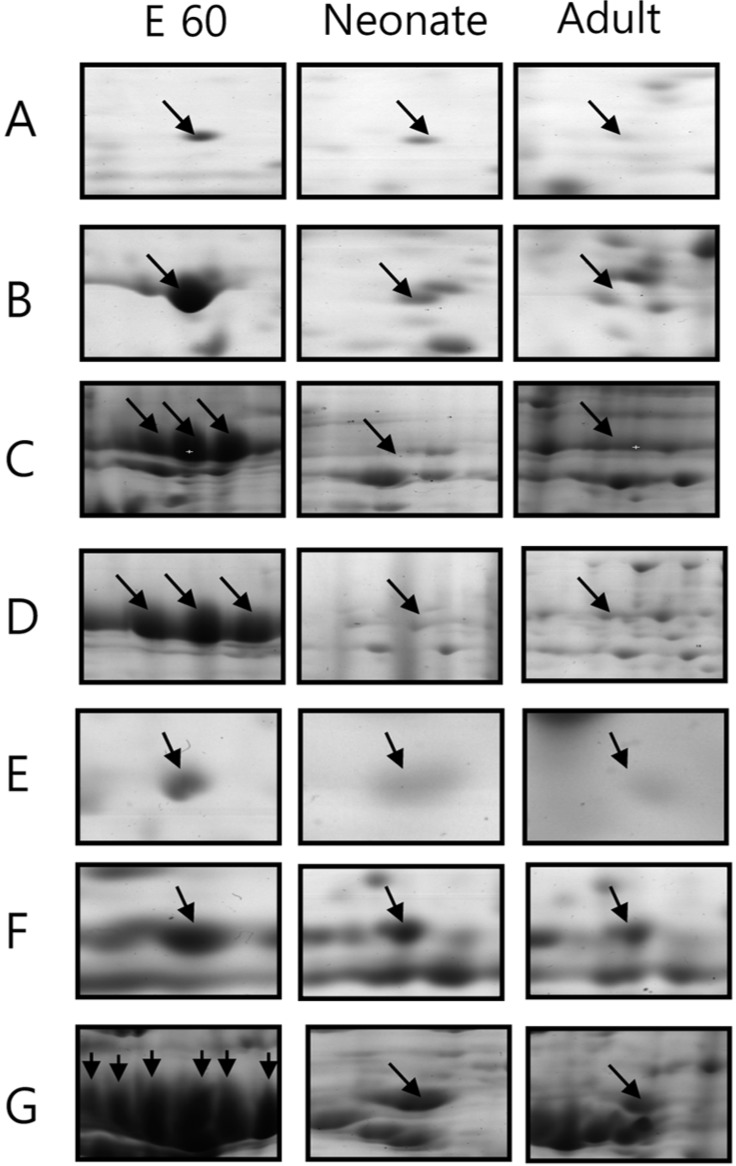
Expression of 7 spots was significantly increased in fetal pigs compared to neonate and adult stages (Figure 4). Apolipoprotein precursor, alpha-fetoprotein, transferring and alpha-1-antitrypsin precursor were strong markers of developing pancreas. They were difficult to detect in the developed pancreas.
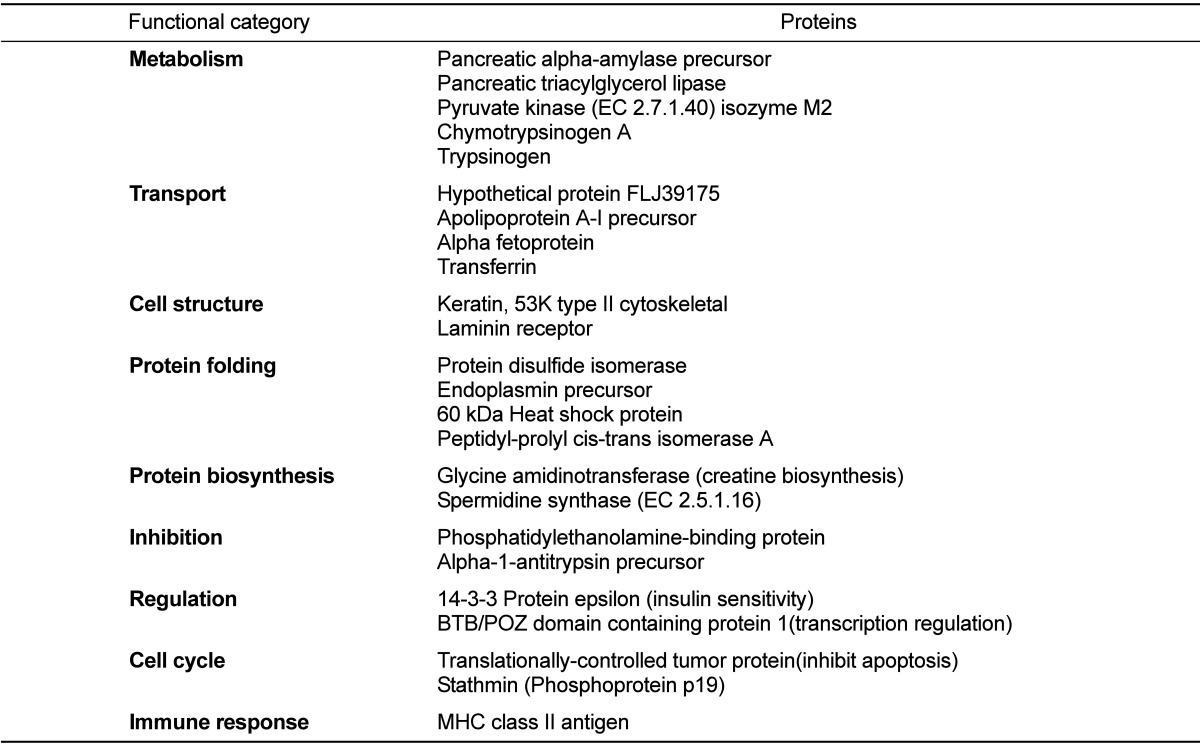
Classification by function
The 24 identified proteins were classified by their functions based on a search at http://harvester.embl.de (Table 2). The functions included glucose or lipid metabolism (n=5), transport of ions like Fe and Ni, or lipid (n=4), maintenance of intracellular structure and cell-to-cell adhesion (n=2), protein folding (n=4), protein synthesis (n=2), anti-proteases (n=2), regulators of signal transduction or insulin sensitivity (n=2), inhibit of apoptosis and induce proliferation (n=2) and immune system related (n=1).
Discussion
Pancreas obtained prior to birth (E60), at the end of pregnancy (neonate) and at adulthood (6 months of age) were analyzed to reveal proteins that were differentially expressed during pig pancreas development. Using 2D proteomics, proteins were resolved and were identified using MALDI-TOF and MASCOT database. We identified 462, 402 and 557 proteins at E60, neonate and 6-month-old adult pig, respectively. Among those identified proteins, 24 proteins showed pronounced differences in protein expression level at each developmental stage. The fetus and the neonatal pig pancreas had similar protein expression patterns on 2-DE gels, while the adult pig pancreas had a significantly different pattern. In the adult pig, digestive enzymes, such as pancreatic alpha-amylase precursor, trypsin precursor, pancreatic triacylglycerol lipase, protein disulfide-isomerase precursor, endoplasmin precursor and glycine amidinotransferase, which are associated with protein synthesis and formation of tertiary structure, consisted most of the dramatically up-regulated proteins on the 2-DE map.
Comparing protein expression levels in the E60 and the neonatal pig (Figure 1), proteins in the fetal pig were more acidic and high in molecular weights. These included stathmin, apolipoprotein A-I precursor, alpha-fetoprotein, transferrin, peptidyl-prolyl cis-trans isomerase A (cyclophilin A) and alpha-1-antitrypsin precursor. These proteins gradually decreased during pancreas development. Stathmin is the tubulin-binding protein but a neuroproteomic search identified it as a novel Cdk5 substrate. Stathmin is phosphorylated by Cdk5 [21] and is involved in cancer development [22,23]. Stathmin gene overexpression is important in the maintenance of the malignant phenotype in tumor cells, and the blocking efficacy and tumor specificity of this target has been addressed in clinical trials [24]. Small interfering RNA-induced repression of stathmin decreases cell proliferation, viability and clonogenicity in mutant p53 tumor cell lines [25]. Although the relationship between stathmin and cell development is unclear, since stathmin is associated with aberrant cell proliferation, it is expected that stathmin contributes to cell development in some manner.
Apolipoprotein A-I (apoA-I) is the major lipoprotein component of high-density lipoprotein, and plays an important role in reverse cholesterol transport [26]. It also is an anti-inflammatory and anti-atherosclerosis molecule and functions as a potent inhibitor of dendritic cell differentiation and maturation [27]. Apo A-I has two major sites of synthesis: the intestine and the liver. Apo A-I is expressed in the pancreas and 12% of the liver [28]. Presently, its expression decreased gradually according to the developmental stage, which may be due to lack of cell type specification during early development because liver and pancreas have the same developmental origin. In addition, the liver controls lipid metabolism and repression of inflammatory response, which may contribute to the development of the pancreas (although there is scant evidence in support of this).
The liver and yolk sac of fetuses the principle sites of alpha-fetoprotein (AFP) synthesis in ontogenesis, and the dynamics of AFP in ontogenesis from the early embryonic period through mid-pregnancy to pregnancy termination to AFP shut-down in early postnatal period [29]. AFP plays an important role in regulation of immune system, which is associated with repression on the immune system, such as decrease in natural killer cell activity [30], induction of T suppressor cells [31,32], repression of mitogenic response of PHA and Con A [33,34,35,36], restriction on T cell differentiation to Ia determinants [37], decrease in phagocytic activity of macrophage [38,39] and decrease in macrophage Ia expression level [40,41,42].
Transferrin, a 76-80 kDa glycoprotein, is responsible for the transport of iron to cells within both the fetal and maternal systems [43]. It is not only the transporter of iron but also has activities of growth factor and repressing reactive oxygen species production [44]. The concentration of AFP, alpha 1-antitrypsin and transferrin in serum increases with age until 40-50 days of gestation and then decreases progressively at birth in pigs [45].
Cyclophilin A (CyP-A) is a soluble cytoplasmic immunophilin involved in T cell differentiation and proliferation. A role for CyP-A in neuronal differentiation has been implied [46]. Alpha-1-antitrypsin (α1-AT) acts as a suicide inhibitor of a wide range of serine proteases. In normal humans, more than 2 g of α1-AT protein is synthesized daily, resulting in a serum concentration of about 2 mg/mL. The primary function of α1-AT protein is inhibition of neutrophil elastase, which may protect from excessive tissue damage [47]. In addition to its well established role of neutralizing elastase activity, in cell lines of both mesenchymal and epithelial origin, α1-AT can enhance DNA synthesis and cell proliferation particularly when insulin is present [46]. The highest amounts of α1-AT were clearly present in the fetal sheep liver, regardless of the stage of gestation. The decline in hepatic α1-AT in term and neonatal animals occurs at a time when the plasma levels of glucocorticoids markedly suggesting that the natural increase in cortisol during late gestation in the sheep fetus may be responsible for this decline in the abundance of hepatic alpha-1-antitrypsin. It may influence the ability of fetus to control proteinases at the fetal maternal interface and influence events leading to parturition, and/or the ultimate demise of the placenta [49].
Disruptions in pancreatic functions cause indigestion problems and altered glucose homeostasis, which can prelude the development of diabetes mellitus, pancreatic cancer and pancreatitis. The pig pancreas is an important resource of xenotransplantation for human diabetic patients, but little is known about its developmental stage at the molecular level. We presented proteomic alterations in pig pancreas by analyzing the changes in protein expression level during development. This study will provide basis for rudimentary understanding development of pig pancreas at the molecular level.
 XML Download
XML Download