

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Ginseng, the root of Panax ginseng C.A. Mayer, and its preparation, red ginseng, are common traditional medicines in Asian countries such as Korea, China, and Japan [1,2]. Red ginseng, which contains many functional chemicals and complexes, is manufactured through techniques utilizing repetitive steaming and drying cycles [3]. This processing results in the formation of additional beneficial compounds, known as ginsenosides, with various pharmacological effects [4] on immune response [5], diabetes [6,7], and Alzheimer's disease [8,9]. The ginsenosides of Panax ginseng are a group of steroidal saponins; over 50 of the compounds have been identified [10], which are generally divided into 2 classes: the protopanaxadiols, with study compounds Rb1, Rb2, Rb3, 20(S)-Rg3, 20(R)-Rg3, and Rd; and the protopanaxatriols, with study compounds Rg1, Re, Rf, Rg5, and Rk1.
Metabolic disorders, including obesity, are chronic worldwide, leading to various detrimental conditions in adults, such as hyperglycemia, dyslipidemia, and hypertension [11,12]. In particular, excessive dietary fatty acids and genetic causes of obesity induce increases in triglyceride (TG) accumulation in fat tissue and the reuptake of free fatty acids into the circulatory system, which contributes to TG accumulation in the liver through fatty acid binding and the action of transport proteins [13,14,15]. Effective control of blood glucose and lipid levels is necessary to regulate or prevent complications such as hyperglycemia and fatty liver [16,17].
Several studies have reported effects of fermentation and enzyme processing on ginseng (including red ginseng) and the development of ginsenosides [3,18,19,20]. Studies have also examined the effects of the resulting ginsenosides in various disease models such as metabolic disorders [21,22,23,24,25], cancer [26,27], immune response [1,28], skin care [29,30], pulmonary system [31], brain injury [32], depression [33], and Alzheimer's disease [34] in both in vitro and in vivo models [35]. Although many pharmaceutical effects of red ginseng and its fermented products have been postulated, the full range of applications of purified enzymes still remains unclear. Indeed, to date, the effects of commercial pectin lyase-modified red ginseng extracts on high fat diet-fed obese mice have not been reported.
Microbial pectin lyase is commonly used for its beneficial effect on various plant products such as carrot [36], and date syrup [37] in the production of functional compounds, but it has not been fully applied to ginseng. In this study, we enzyme-modified red ginseng extracts with microbial pectin lyase and observed its bioavailability in and effects on high fat diet-fed obese mice.
Materials and Methods
Reagents
Dexamethasone, 3-isobutyl-1-methylxanthine (IBMX), insulin, formaldehyde, and Oil Red O solution were purchased from Sigma-Aldrich (MO, USA). Cell culture media, fetal bovine serum, and supplements were obtained from Thermo Scientific (UT, USA)
Sample preparation
A dried mixture of whole red ginseng and red ginseng tail root (4:6 w/w) was extracted 3 times with 70% ethanol. The extracts were then concentrated to 6 Brix with a vacuum evaporator at 40℃. The extracts were incubated with 10% pectin lyase (EC 4.2.2.10, Novozyme, #33095, Bagsvaerd, Denmark), which was prepared from Aspergillus aculeatus and Aspergillus niger cultivates, at 50℃ for 5 days in a shaking incubator (150 rpm). To terminate the reaction, processed extracts were heated at 95℃ for 10 min, and then freeze-dried. The dried GS-E3D consisted of 120.2 mg/g crude saponin containing the following ginsenosides: 5.9 mg/g Rg1, 30.2 mg/g Rb1, 17.6 mg/g Rb2, 2.5 mg/g Rb3, 1.3 mg/g 20(S)-Rg3, 1.4 mg/g 20(R)-Rg3, 27.7 mg/g Rd, 12.6 mg/g Re, 1.5 mg/g Rg5, 0.8 mg/g Rk1, and 4.7 mg/g Rf (International Ginseng and Herb Research Institute, Geumsan, Korea).
Induction and quantification of adipogenesis
To induce adipogenesis, 3T3-L1 preadipocytes were seeded into a 6-well plate at 2×105 cells per well and cultured until fully confluent. After 2 days in the medium, cells were incubated with a differentiation medium containing 1 µg/mL insulin, 1 µM dexamethasone, and 0.5 mM IBMX in standard culture medium for 2 days, followed by additional 48 h with standard culture medium containing insulin alone. The differentiation-maintenance medium was changed every 2 days. To investigate the effect of GS-E3D on differentiation in 3T3-L1, GS-E3D (at 0.1, 0.5, 1.0, 2.0, and 5.0 mg/mL) was added to the differentiation-induction medium. The assay protocol was conducted according to the colorimetric adipogenesis assay kit (Cayman Chemical Company, MI, USA). The absorbance was measured at 500 nm using a spectrophotometer (Infinite 200, Tecan Co, Männedorf, Switzerland) with 96-well plates. Lipid contents were calculated as a percent of control cells.
Animal experiments
Specific-pathogen-free 6-week-old male C57BL/6J mice (18-23 g) were purchased from SAMTAKO Co. Ltd. (Osan, South Korea). The experiments were performed in accordance with the principles of and approval from the Ethics Committee of the Wonkwang University, Iksan, Korea (WKU12-47). All animals, which were maintained in a temperature-controlled room (temperature 22±2℃, humidity 50±5%) with a 12/12 h light/dark cycle, were acclimatized to the laboratory environment while housed in individual cages for 1 week before the experiment. Obesity was induced by a 60% fat calorie diet (high fat diet, or HFD), and mice were randomly divided into 5 groups (10 mice per group) after the first 3 weeks on high fat diet (AIN-93G, cat., #101556; Research Diet, Inc., NJ, USA). The study groups included: the normal diet group; the high fat diet-vehicle group; high fat diet+0.1 g/kg GS-E3D (0.1-GS-E3D); high fat diet+0.3 g/kg GS-E3D (0.3-GS-E3D); and high fat diet+1.0 g/kg GS-E3D (1.0-GS-E3D). Depending on the assigned group, placebo (water) or 3 doses of GS-E3D (0.1, 0.3, or 1.0 g/kg) were administered orally, twice daily for 4 weeks. At the end of the experiment, 4 h-fasted mice were anaesthetized using a mixture of xylazine/ketamine (1:3, v/v), and blood samples were collected and stored at -80℃ until their analyses (Figure 1).

Measurements of glucose, triglyceride, cholesterol, and leptin levels
After 4 weeks of treatment with and without GS-E3D, blood samples were collected and centrifuged at 3,000 rpm for 15 min at 4℃, and then levels of serum glucose, total cholesterol (T-CHO), high-density lipoprotein cholesterol (HDL-C), low-density lipoprotein cholesterol (LDL-C), triglyceride (TG), and leptin were measured by Green Cross Reference Lab (Yongin, Korea).
Measurements of body, organ, and fat weight
Body weight was measured once a week during the feeding period. Internal organs were dissected and weighed. All body, organ and fat was weighed to the nearest 0.01 g using an electronic balance (Sartorius, Göttingen, Germany). Fat tissue samples also were dissected, weighed, and then processed for further histological experiments.
Histological analysis
Histological analysis was conducted following routine methods. Briefly, the dissected epididymal adipose tissues were fixed in 10% neutral buffered formalin for histological analysis and embedded in paraffin. The paraffin sections were cut at a thickness of 4 µm and stained with hematoxylin and eosin (H&E). Adipocyte sizes were measured in randomly chosen microscopic areas from individual animals using a Nikon microscope system (TE-2000, Tokyo, Japan). Average adipocyte size was calculated by dividing the each chosen microscopic area by total adipocyte cells in the area [38].
Statistical analysis
All data were expressed as mean±standard error of the mean (SEM), and differences between groups were analyzed using one-way ANOVA (Duncan's multiple-range test). The effects of GS-E3D on body weight, food intake and water intake in experimental period were compared by two-way repeated measures analysis of variance (ANOVA) with Bonferroni's post-test analysis. All analyses were performed using SPSS 10.0 (SPSS Inc. USA). Each value was the mean of at least 3 separate experiments in each group. Statistical significance was set at P<0.05.
Go to : 
Results
Cytotoxicity of GS-E3D on 3T3-L1 adipocyte cells
To determine the cytotoxic effects of GS-E3D on 3T3-L1 adipocyte cells, we performed a tetrazolium (MTT) assay. Treatment of GS-E3D did not inhibit 3T3-L1 cell viability at 5 mg/mL (>95%), but exposure of 10 mg/mL GS-E3D to cells showed high growth inhibition at 48 h (approximately<40%) (Figure 2). Therefore, we did not use a dose of 10 mg/mL GS-E3D for further experiments.
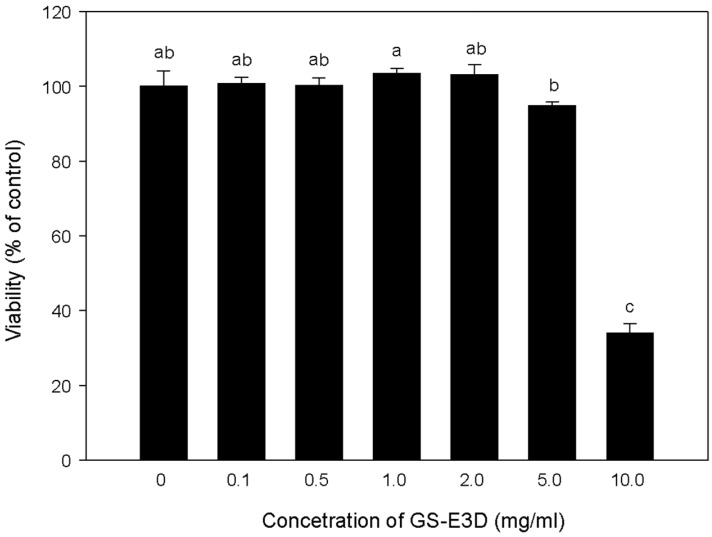 | Figure 2Cell viability of 3T3-L1 adipocytes against pectin lyase-modified red ginseng extracts (GS-E3D). 3T3-L1 cells (5×103 cells/well) were grown in 96-well plates and treated with different concentrations (0.1, 0.5, 1, 2 and 5 mg/mL) of GS-E3D. Differences were tested for significance by a one-way ANOVA (Duncan's multiple-range test). a,b,cValues in the row with different superscript letters are significantly different; P<0.05.
|
GS-E3D reduces adipocyte differentiation in 3T3-L1 cells
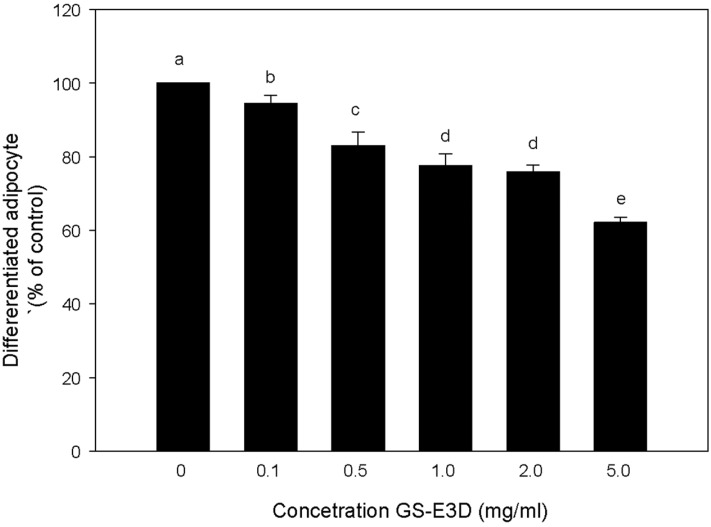
A 3T3-L1 differentiation assay was performed to investigate the effect of GS-E3D in 3T3-L1 cells using an Oil Red O staining-based assay kit (Cayman Chemical Company, MI, USA). Differentiated 3T3-L1 cells were treated every 48 h with GS-E3D at the indicated concentration, for 5 days. Based on quantification of lipid content, results showed that GS-E3D treatment at 0.1, 0.5, 1.0, 2.0 and 5.0 mg/mL in 3T3-L1 cells could inhibit lipid accumulation, as compared to untreated cells, in a dose-dependent manner (Figure 3). At 5.0 mg/mL, GS-E3D treatment showed a maximum inhibitory effect without cytotoxicity (Figure 2). These results suggest that GS-E3D may reduce adipogenesis and lipid accumulation in fat tissues.
Effects of GS-E3D on body weight, food intake, and water intake
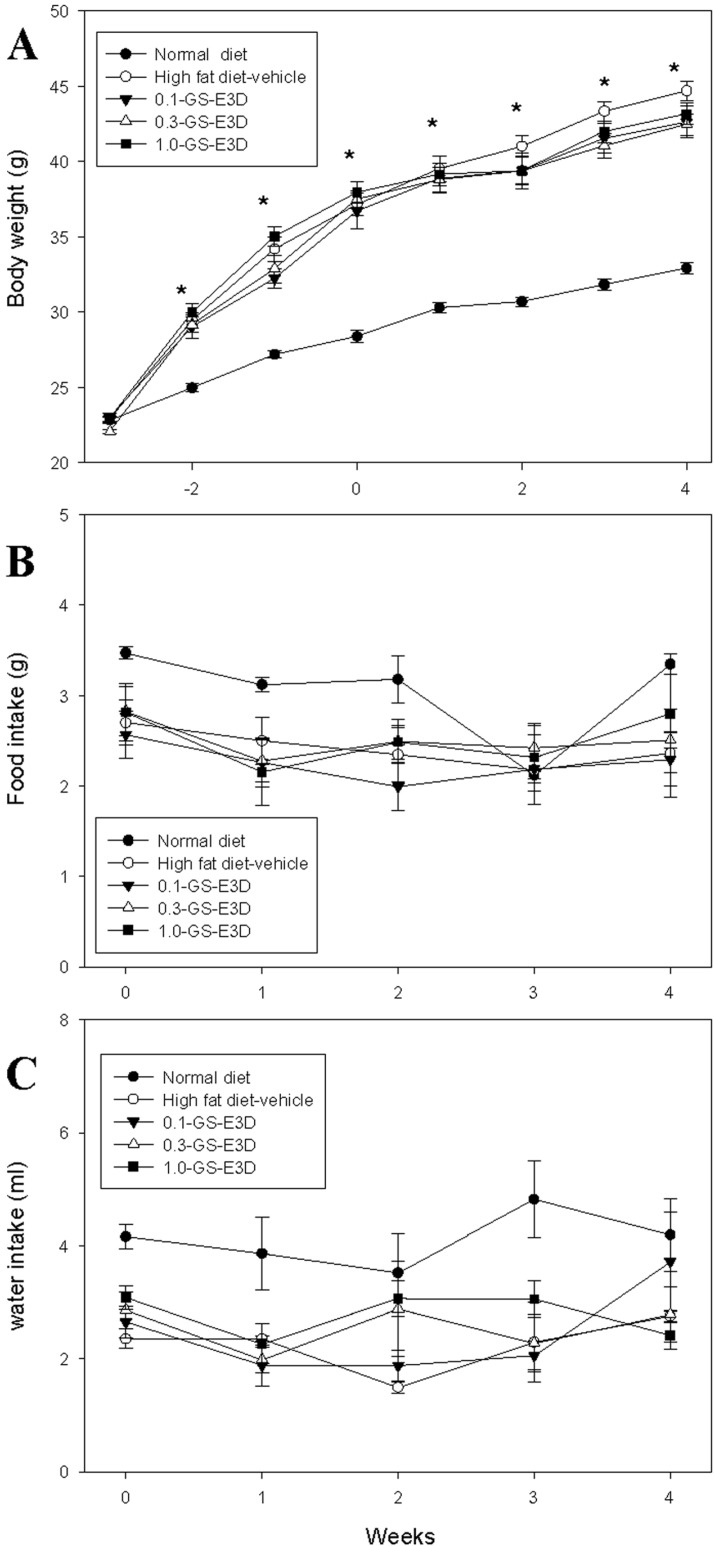
The effects of GS-E3D on body weights are shown in Figure 4A. After 4 weeks of treatment with GS-E3D, the body weight of the high fat diet-vehicle group was significantly higher than that of the normal group. In contrast, the body weights of the GS-E3D groups were significantly lower, compared to the high fat diet-vehicle group. Although the body weight was significantly reduced in the GS-E3D group at 4 weeks after supplementation, there were no significant observed differences in body weight from 1 to 3 weeks. However, food intake and water intake did not significantly differ between each group (Figures 4B and C).
Effects of GS-E3D on blood glucose in high fat diet-induced obesity model mice
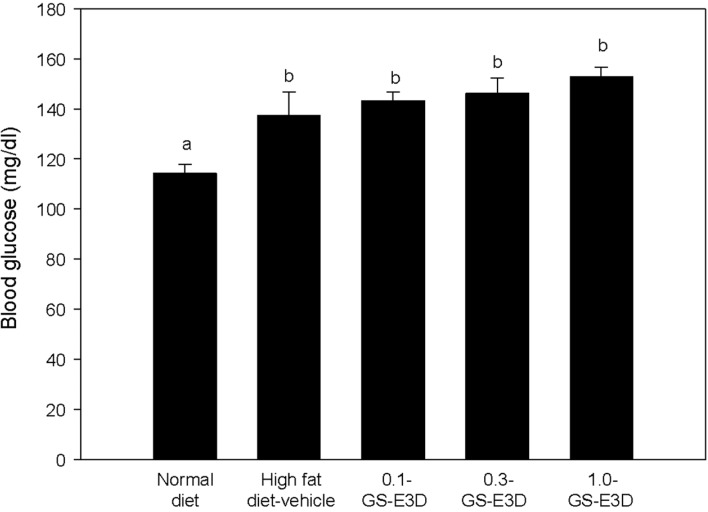
Figure 5 shows the effects of GS-E3D on blood glucose levels after 4 weeks. Blood glucose levels were measured at 4 weeks after twice-daily administration. Compared to the normal group, the high fat diet group had higher blood glucose levels. However, there were no significant changes in glucose levels by GS-E3D treatment.
Effects of GS-E3D on total cholesterol, LDL and HDL cholesterol, triglyceride, and leptin in high fat diet-induced obesity model mice
Lipid levels (T-CHO, LDL-C, HDL-C, and TG) of experimental mice were analyzed (Figure 6). The control (high fat diet-fed mice) group showed high levels of T-CHO, LDL-C, and HDL-C when compared to the regular chow diet group (Figure 6A-C), but no significant increase was observed in TG concentration (Figure 6D). As shown Figure 6, T-CHO, LDL-C, HDL-C, and TG levels were significantly reduced in the GS-E3D 0.5 and GS-E3D 1.0 groups, compared to the control group. These results suggest that GS-E3D improved blood lipid profiles.
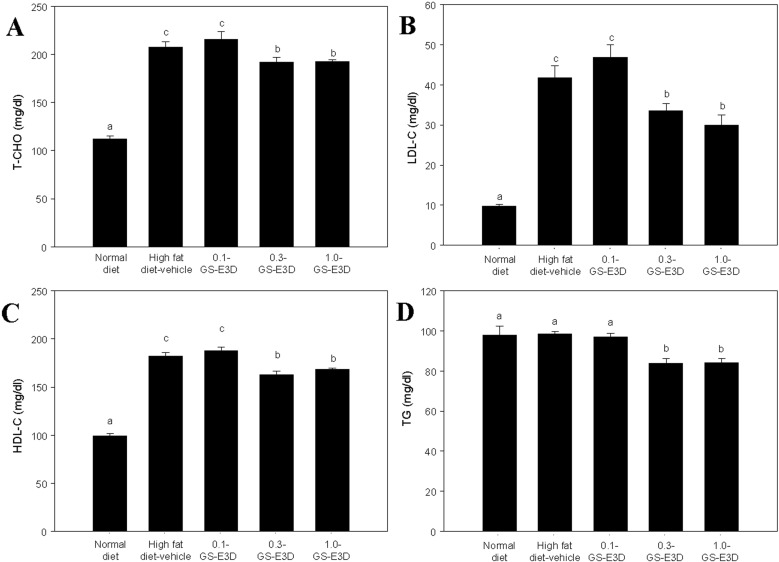 | Figure 6Effect of GS-E3D on changes in blood parameters in an obesity model induced by a high-fat diet. (A) total cholesterol (T-CHO), (B) low density lipoprotein (LDL-C) and (C) high density lipoprotein (HDL-C), (D) triglyceride (TG) in the whole blood of mice. a,b,cValues in the row with different superscripts are significantly different; P<0.05. Data are shown as the mean±SE (n=7).
|
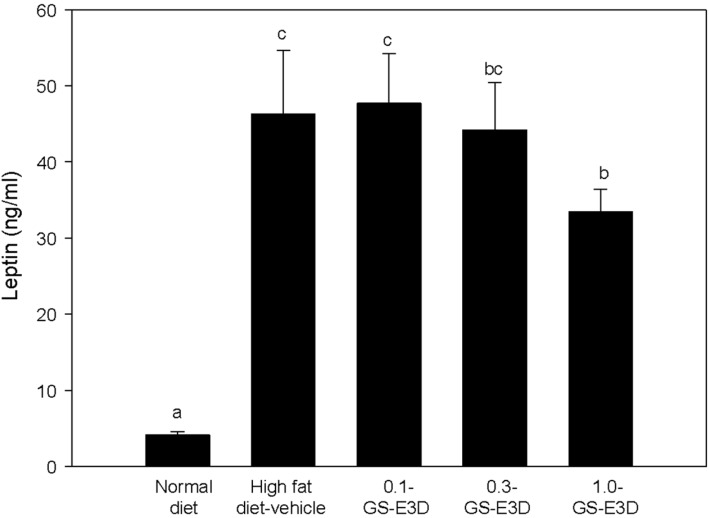
Effects of GS-E3D on leptin level in high fat dietinduced obesity model mice
The results of leptin levels are shown in Figure 7. The serum leptin levels were significantly elevated in high fat diet-induced obese mice. In contrast, oral administration of GS-E3D significantly reduced serum leptin levels in a dose-dependent manner, compared to the control group.
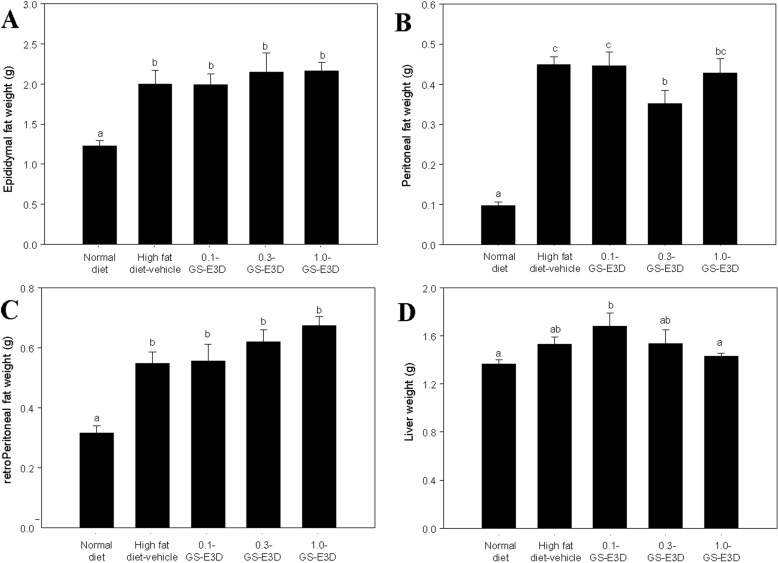
Effects of GS-E3D on fat tissue and liver weight in high fat diet-induced obesity model mice
As shown in Figure 8A-C, fat weights were significantly higher in the high fat diet-induced obese mice group than in the normal group. However, there were no significant differences in the fat weights between the control and GS-E3D groups. Although there was no effect of GS-E3D on fat weights, administration of GS-E3D reduced liver weight overall (Figure 8D). The liver weight of high fat diet-induced obese mice was significantly higher than that of the normal group. In contrast, the GS-E3D 1.0 group showed significantly reduced liver weight; however, the GS-E3D 0.1 and GS-E3D 0.3 groups showed no significant differences.
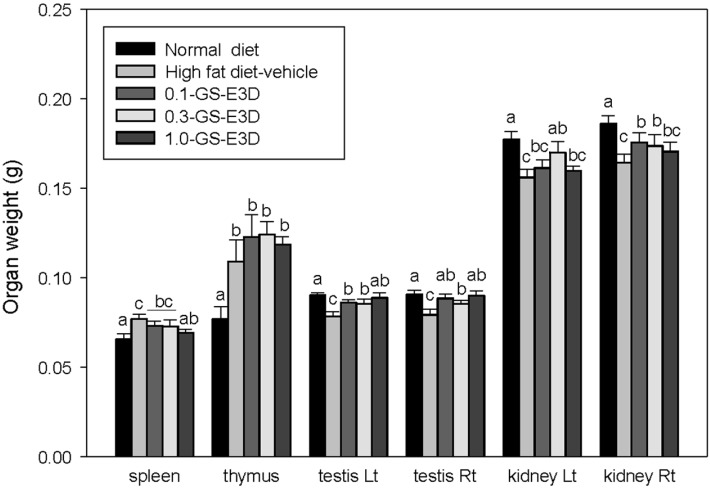
Effects of GS-E3D on organ weight in high fat diet-induced obesity model mice
Figure 9 shows the effects on organ weight for each group. The organ weights were measured at 4 weeks. Significant increases in the weights of spleen and thymus were observed, but the weights of testis and kidney decreased on the high fat diet. In contrast, the administration of GS-E3Ds reduced the weights of spleen and increased the weight of testis and kidney in high fat diet-induced obese mice. Although the weight of thymus in both the control and GS-E3D groups were higher than the normal group, these phenomena were induced through reduction of immune function [39,40]. These results suggest that GS-E3D regulates the innate function of organs involved in metabolism and immunity.
Effects of GS-E3D on adipocyte size in high fat diet-induced obesity model mice
The effects of GS-E3D on changes in adipocyte size in high fat diet obese mice are shown in Figure 10. High fat diet obese mice showed significantly larger adipocytes from epididymal adipose tissue (Figure 10B), but administration of GS-E3D markedly reduced the size of adipocytes, in a dose-dependent manner.
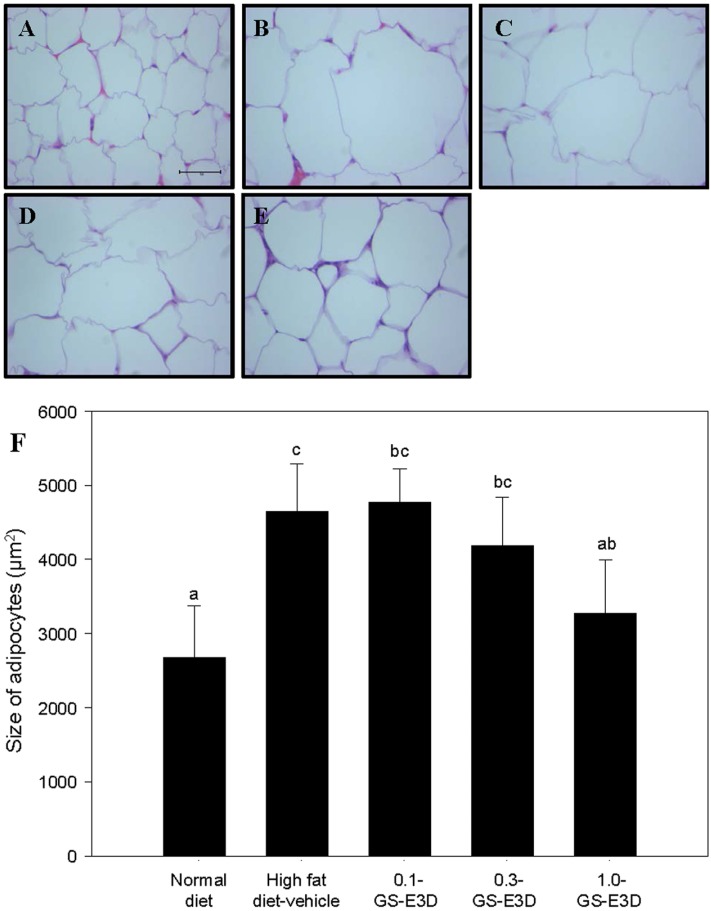 | Figure 10Effects of GS-E3D on histological adipocyte size in a mouse obesity model induced by a high-fat diet. (A) Normal group, (B) high fat diet-vehicle group, (C) high fat diet+ GS-E3D 0.1 g/kg, (D) high fat diet+GS-E3D 0.3 g/kg, (E) high fat diet+GS-E3D 1.0 g/kg, (F) quantitative analysis (400× magnification). a,b,c,dValues in the row with different superscript letters are significantly different; P<0.05. Data are shown as mean±SE (n=7).
|
Effects of GS-E3D on hepatic steatosis in high fat diet-induced obesity model mice
To determine ameliorative effects of GS-E3D on high fat diet-induced hepatic steatosis, we observed lipid droplet contents through an H&E stain in liver after 4 weeks (Figure 11). High fat diet mice had significantly increased steatosis scores, but administration of F-BS reduced the number of lipid droplets in a dose-dependent manner (Figure 11F).
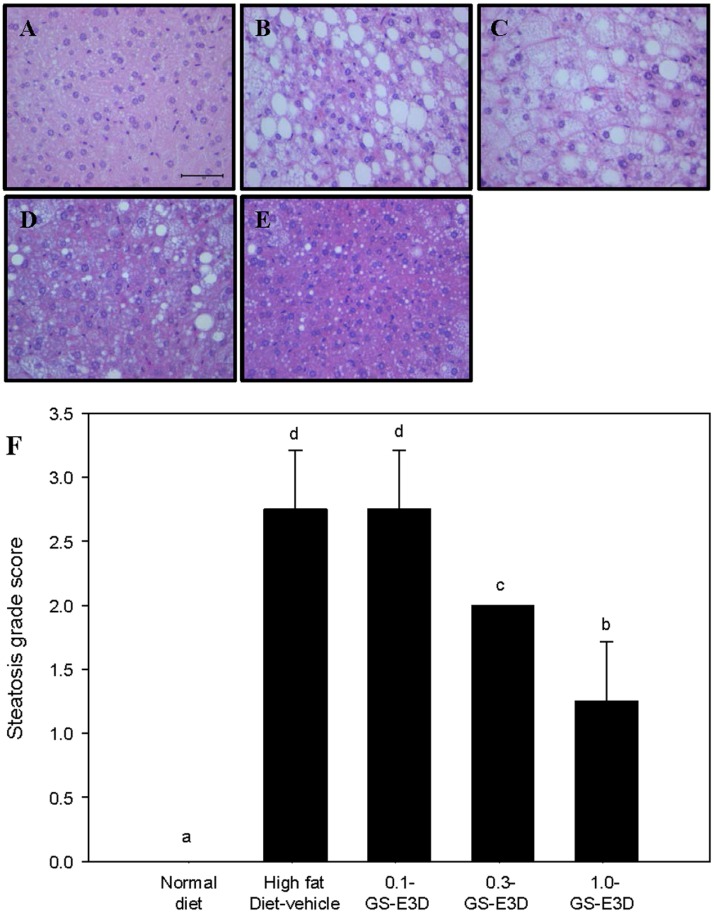 | Figure 11Effects of GS-E3D on histological hepatic steatosis in a mouse obesity model induced by a high-fat diet. (A) Normal group, (B) high fat diet-vehicle group, (C) high fat diet+GS-E3D 0.1 g/kg, (D) high fat diet+GS-E3D 0.3 g/kg, (E) high fat diet+GS-E3D 1.0 g/kg, (F) steatosis grade score. (400× magnification). Scale bar=100 µµm. a,b,c,dValues in the row with different superscript letters are significantly different; P<0.05. Data are shown as mean±SE (n=8).
|
Go to :
Discussion
Ginseng has been widely recognized for several thousand years as a valuable plant and useful medicine in the Asia-Pacific region, and in many countries such as Korea, China, Japan, and even Canada [7,25]. Typically, orally-administrated ginseng is broken down into various molecules, including ginsenosides, through exposure to gastric juices, digestive enzymes, and intestinal bacteria. Consequently, the absorption efficacy of ginseng's components may differ due to the varying resident microenvironments in the individuals consuming it [41]. Several groups have examined microflora in conjunction with enzymes in several studies involving red ginseng, with the aim of increasing efficacy [22,34,35,26,30,32]. Similarly, GS-E3D is produced from red ginseng extracts that were enzymatically modified with commercial pectin lyase (EC 4.2.2.10); however, in this study, we use tail root as a source of red ginseng and do not report on these methods.
Our aim was to investigate the effects and bioavailability of GS-E3D on high fat diet-fed obese mice. In addition, to develop a novel processing procedure of red ginseng extracts, we increased the amount of red ginseng tail root to exceed the amount of whole red ginseng (>1.5 fold, w/w) in extracts, as tail root contains more ginsenosides in red ginseng. In other hands, we should compared the effects of red ginseng extracts with G3-E3D but not observed significant differences between both extracts in experimental model as expected, unfortunately. However, GS-E3D and its processing procedures will helpful for absorption and utilization of effective molecules of red ginseng in other models.
We also observed ameliorative effects of GS-E3D on hepatic steatosis (Figure 11) and demonstrated histologic data with statistics. GS-E3D was significantly reduced lipid droplets in liver tissues but not recovered completely other phenomena such as observe of eosinophilic cytoplasm in H&E stained tissue sections. Indeed, lower food intake were observed in normal diet group at 3 weeks after first administration of GS-E3D or control, but it was unidentifiable things at the specific time couse. Therefore, these apparent symptom must be recognized and need further study.
In the present study, we found beneficial effects of GS-E3D for high fat diet-fed obese mice. Administration of GS-E3D resulted in improved blood glucose levels, lipid profiles, and leptin levels in high fat diet-fed obese mice. Moreover, GS-E3D reversed the progression of fatty liver by high fat diet-fed mice in the same animal model. Indeed, GS-E3D administration was found to be effective in reducing adipogenesis and lipid metabolism in high fat diet-induced obesity and liver steatosis models. The treatment decreased adipocyte differentiation, and ameliorated blood lipid parameters during a 4-week trial. Treatment significantly reduced the size of epididymal adipose tissue and conclusively ameliorated high fat diet-induced hepatic steatosis. These results suggest that GS-E3D as a dietary supplement may have beneficial effects on obese individuals, and may be useful as a health-care product. In the future, studies on the ginsenoside composition and the detailed mechanisms of processing techniques of red ginseng are required.
Go to :
 XML Download
XML Download