

 PDF
PDF ePub
ePub Citation
Citation Print
Print


The increasing shortage of human donor organs has provoked interest in the possible use of animal organs for transplantation into humans [1,2,3]. Organ transplants from nonhuman primates have been studied as replacements for human organs. However, using organs of nonhuman primates can be dangerous, and their organs can be too small for transplanatation into adult humans [1]. Pigs are the most likely candidate for xenotransplantation [4,5]. Moreover, as a potential xenotransplantation source, pigs have to be managed under conditions of stringent bioexclusion. Miniature pigs produced by a selective breeding system are recommended [6,7].
Miniature pigs have many physiological similarities with humans and provide some breeding and manipulation advantages compared to non-human primates [8]. The organs commonly used from miniature pigs are the heart, kidney, liver and pancreas. The pancreas controls endocrine, exocrine and glucose metabolism through the creation of insulin. Porcine pancreatic tissue can be a good resource of transplantation therapy for human diabetic patients [9,10,11]. Transplantation of porcine pancreatic islets into non-human primates or other species has been documented [12,13,14]. However, the characteristics of pancreatic protein expression in pigs are ill-understood, while the expression of pancreatic proteins in humans and rodents is better defined.
Presently, pancreatic protein expression was examined chronologically using a miniature neonate pig (4-day-old), miniature piglet (19-day-old) and miniature adult pig (14-months-old). Proteins that were differentially expressed during development were revealed by two-dimensional electrophoresis (2-DE) and matrix assisted laser desorption/ionization-time of flight mass spectrometry (MALDI-TOF MS). While the repository of identified porcine proteins in Swiss-Prot and NCBI databases are relatively scant, these sources were used presently to help identify the differentially expressed proteins. The data should aid future proteome analyses.
Materials and Methods
Laboratory animals
The miniature pigs were housed in the air barrier facility of the Center for Animal Resource Development at Seoul National University. Three pigs (male 4-old-, 19-day- and 14-month-old miniature pig) having different ages and developmental stages were used. They were conditioned in the same room at 24±2℃ with a 12/12 (light/dark) cycle. Air and water were filtered, and all equipment and food were sterilized before use. This study was approved as an animal use protocol by the Institutional Animal Care and Use Committee at SNU (SNU-IACUC approval number: SNU-060613-5). Intramuscular anesthesia was achieved using a mixture of 2 mL ketamine · HCl (50 mg/mL/kg), 1 mL xylazin (2.3 mg/mL/kg) and 1 mL atropine sulfate (0.5 mg/mL/10 kg) during every painful/stressful procedure. The abdomen of each pig was shaved and disinfected with polyvinylpyrrolidone-iodine. The animals were immobilized on a surgical plane-table by adhesive ties. A median laparotomy was performed. The pancreas from the animals were nipped off aseptically and stored in liquid nitrogen while being transported to the laboratory.
Sample preparation for 2DE
The pancreas of each pig (one miniature pig for each age stage, respectively)was removed immediately after sacrifice and stored at -70℃ until used. In briefly, the peritoneal of the miniature pigs under the anesthesia were pre-chilled by filling of large quantity of ice before removing the pancreas. The pancreas was collected from the distal end of normal pancreas immediately from the animals and snap-frozen in liquid nitrogen, and stored at stored at -70℃. The frozen pancreas tissues (1 g) in a mortar were grounded to powder under the constant addition of liquid nitrogen. The powder is normally suspended in 900 µL lysis buffer (7 M urea, 2M thiourea, 2% w/v CHAPS, 2% pharmalyte pH3-10, 100 mM DTE). Samples were centrifuged at 50,000 rpm at 4℃ for 1 h. The supernatant was carefully removed and immediately frozen at -70℃.
2DE
2D-polyacrylamide gel electrophoresis (PAGE) was performed as previously described [15]. Aliquots containing 1 mg total protein were diluted in lysis buffer to a total volume of 450 µL. The samples were applied to a 240 mm, immobilized, nonlinear pH 3-10 IPG Drystrip (Amersham Pharmacia Biotech, Piscataway, NJ, USA), which was rehydrated for at least 12 h. After rehydration, the strips were focused at 30 V for 3 h, 100 V for 1 h, 200 V for 1 h, 500 V for 1 h, 1,000 V for 1 h and finally at 8,000 V for 11 h to obtain approximately 90,000 Vh (Ettan™ IPGphor™ II IEF systems; Amersham Pharmacia Biotech). Once isoelectric focusing was completed, the strips were equilibrated in 6M urea containing 20% v/v glycerol, 2% w/v sodium dodecyl sulfate (SDS) and 0.01% w/v BPB with 10 mM tributyl phosphine (Flukachemie, Buchs, Switzerland). SDS-PAGE was performed using an 8-18% separating gel without a stacking gel using the Ettan Dalt system (Amersham Pharmacia Biotech) twice per each aged-group for the confirmation of separated gel-spot trends. The second dimension electrophoresis was carried out overnight at 3 W/gel at 20℃. The gels were stained with Coomassie G-250 (Bio-Rad Laboratories, Hercules, CA, USA).
Protein visualization and image analysis
The gels were stained with Coomassie G-250 (Bio-Rad) as previously described [12]. The stained gels were scanned using a GS 800 photometer (Bio-Rad) and analyzed with the ImageMaster™ 2D Platinum Software version 5.0 (GeneBio, Geneva, Switzerland). The digitalized 2DE gel images were compared by a matching method (Image master 5.0; Amersham Biosciences). Differentially expressed spots (>3-fold and <1/3-fold) were analyzed and annotated.
In-gel digestion
The spots were cut into smaller pieces and digested using 12.5 ng/µL trypsin (Promega, Madison, WI, USA) in 50 mM ammonium bicarbonate, pH 8.0, as previously described [16]. For MALDI-TOF MS analysis, the tryptic peptides were concentrated on a POROS 50 R2 column (Applied BioSystems, Foster City, CA, USA). After subsequent washing steps of column with 70% acetonitrile (can) in 5% FA, 100% can and 5% FA, the samples were loaded into a POROS 50 R2 column and washed with 5% FA. The sample was eluted with 2 µL of matrix solution consisting of 10 mg/mL α-cyano-4-hydroxycinnamic acid (Sigma-Aldrich, St. Louis, MO, USA) and then dropped onto a MALDI sample plate [24].
Protein identification by MALDI-TOF
MALDI-TOF MS was performed using a Voyager DE-PRO spectrometer (Applied Biosystems) equipped with a 337 nm nitrogen laser. The instrument was operated at an accelerating voltage of 20 kV, positive ion reflection mode, voltage grid 74.5%, guide wire voltage 0% and delay-time of 120 ns. The spectra were internally calibrated using the trypsin autolysis products (842.51 [M+H] and 2211.11 [M+H]), and searching in Swiss-Prot identified the proteins and NCBI database using Mascot (Matrix Science, London, UK). Monoisotopic peaks [MH+] were selected and all the searches were analyzed with a 50 ppm mass tolerance.
Go to : 
Results
Protein identification by MS
Porcine pancreatic proteins that were differentially expressed were identified and compared according to the developmental stage. Pancreatic proteins were extracted and analyzed by 2DE. After the resolved proteins were visualized by Coomassie blue staining, 2-DE was repeated in three independent experiments for the three pairs of matched samples (Figure 1). For each sample, the three gels with the best resolution were selected for analysis. Spot intensities were analyzed using Image Master™ software (Geneva Bioinformatics, S.A, Geneva, Switzerland). Among the 289 protein spots detected, 13 differentially expressed spots were identified. The protein spots up-regulated more than 3-fold or down-regulated less than 1/3-fold down-regulation were selected, excised from the gels and digested. Peptide fingerprinting was accomplished by MALDI-TOF MS. Peptide mass data were identified by Mascot. Thirteen differentially expressed protein spots were evident. Of these, nine were identified (Table 1) and their functions established based on a search at http://www.ebi.ac.uk/ego (Table 1).
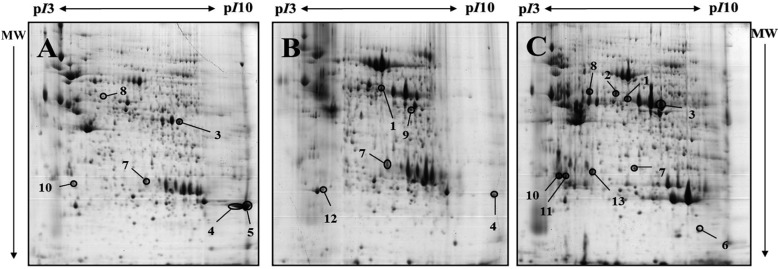 | Figure 1Representative 2-DE gels of pancreas of 4-day-old, 19-day-old and 14-month-old miniature pigs by developmental stages, visualized by Commassie blue staining. Samples of 1mg protein were separated on pH3-10 non-linear IPG strip (24cm) following by 8-18% gradient SDS-PAGE in the second dimension. Proteins were detected by Coomassie brilliant blue G-250 and compared using ImageMaster™ 2D Platinum Software version 5.0. A: 2-DE gel of 4-day old miniature pig(neonate), B: 2-DE gel of 19-day old miniature pig, C: 2-DE gel of 14-month old miniature pig. The closed circles indicate 13 differentially expressed protein spots in the 4-day-old miniature neonate pig, 19-day-old miniature piglet and 14-month-old miniature adult pig.
|
Classification of the identified proteins
The increased and decreased intensities of the selected spots were ascertained chronologically by comparison of the miniature pigs from the neonate stage to adulthood (Figure 2). The expression of pancreatic triacylglycerol lipase (spot 2), tyrosine-rich protein 1 precursor (spot 6) and interleukin-1 receptor type Iprecursor (spot 8) were down- and up-regulated. Although spot 10 displayed a similar intensity, correct identification was not achieved. Pancreatic alpha amylase precursor (spot 3) and high mobility group protein B2 (spot 13) were both up-regulated. Spots 1 and 7 displayed similar intensity patterns, but correct identification was again not achieved. Hemoglobin alpha-subunit spots (spots 4 and 5) were shown side-by-side; the spot intensities were variable, being either both down-regulated or down-regulated and unchanged, respectively. Bisphosphoglyceratemutase (2, 3-bisphosphoglycrerate mutase; spot 11) expression was either unchanged or up-regulated. The expression of alpha-enolase (2-phospho-D-glyceratehydro-lyase; spot 9) and spot no. 12 was up-regulated and down-regulated, respectively. Spot 12, which showed similar spot intensity, was not identified correctly.
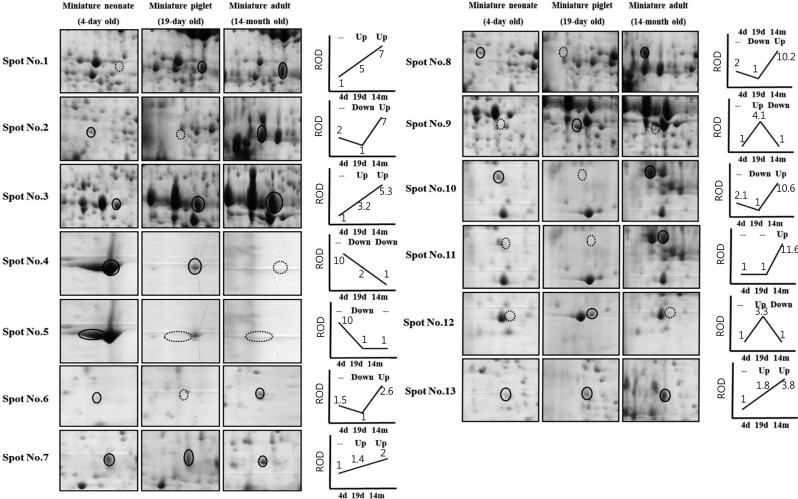 | Figure 2Two-DE images of alterative spots of pancreas according to developmental stage. The circles indicate differentially expressed protein spots and the dot circles show down-expressed patterns. The expression patterns of the different three stages are presented as simple line graphs. Relative optical (ROD) are indicated as relative numbers under the lines compared with the lowest protein optical density (the lowest optical density is made as an '1') of each spot.
|
Go to :
Discussion
This study is, to our knowledge, the first report of protein expression of pancreas in miniature pigs according to the developmental stage. Although xenotransplantation of pancreas using pathogen-free miniature pigs has been explored for a few decades, few studies have examined the proteome of the whole pancreas of miniature pigs at the chronological developmental stages (in this study, neonate, 19-day old and 14-month old miniature pigs). A comprehensive understanding of protein expressional alteration in the pancreas can provide useful biological and clinical information. About 300 proteins with isoelectric points of 3-10 and molecular weight of 10-100 kDa were detected. Of 289 spots, 13 were significantly changed during development; of these, nine spots were functionally identified. Of these identified proteins, few were directly related with pancreas functions. For instance, pancreatic triacylglycerol lipase is required for efficient dietary triglyceride digestion [18,19]. Although this enzyme may play a role in fat metabolism, its expression varies developmentally and by tissue [19]. Pancreatic alpha amylase precursor is found in the pancreas and salivary gland [20]. Alpha-amylase is a typical secretory protein that is subject to glycosylation machinery during its biosynthesis [21,22]. The glycosylation is preceded by proteolytic processing during the biosynthesis of the alpha-amylase molecule [23]. The mammalian interleukin-1 receptor type 1-Toll-like receptor superfamily has important roles in innate immunity, inflammatory responses, stress response and fatal immune disorders [24,25,26,27], and it undergoes intramembrane proteolytic processing [28]. Several studies have reported the relationship of interleukin-1 receptor type 1 with pancreatitis [29,30,31]. In addition, inflammation in type 2 diabetes has been linked to interleukin-1 receptor type 1 [30]. Alpha-enolase is a 48-kDa protein that is crucial in the glycolytic pathway [32], where it catalyzes the formation of phosphoenolpyruvate from 2-phosphoglycerate, the second of the two high energy intermediates that generate ATP in glycolysis [33].
Despite the numerous studies of xenotransplantation between pigs and humans for considerable times, the proteomic alterations of various organs during development, whether or not these changes are critical in the particular developmental stage, has not been considered. Additionally, data concerning the proteome of porcine pancreatic tissue are scant [5], with no attempt to investigate the transplantation-relevant proteomic expressional relationships and differences between human and miniature pigs.
Knowledge of the alteration of the protein complement would provide valuable information to overcome the problems posed by the immunologic barrier, microbiologic differences and xenospecies, which presently hinder xenotransplantation. From the present data, the pancreactic proteomic alterations in the developmental stages appear to contain several enzymatic activities and immune responses, and have hematological relevance. Although the scope of the data was hindered by technical problems, the data provides the first reference of the protein expression during pancreas development, which will be useful in further proteome analyses of the porcine pancreas with respect to xenotransplantation.
Go to :
 XML Download
XML Download