

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Alzheimer's disease (AD) is a common mental disease of dementia showing memory impairment as well as defects with language, decision-making ability, judgment, and personality [1,2]. Although the cause and progression of AD are not fully understood, previous research indicates that this disease is associated with amyloid plaques and neurofibrillary tangles in the brain [3]. Plaques are insoluble deposits of amyloid-beta, which consists of small peptides, 39-43 amino acids in length, formed in the outside regions of cellular material surrounding neurons [4-6]. Further, tangles are formed by aggregation of the microtubule-associated protein Tau after it has become hyperphosphorylated and accumulated in neuronal cells [4].
Although no drug has been shown to inhibit the onset or progression of AD, five pharmaceutical medications are widely applied to treat the cognitive and functional impairments of AD. Four of these are acetylcholinesterase inhibitors, which act to reduce the rate of destruction of acetylcholine (ACh), thereby maintaing the concentration of ACh in the brain and preventing death of cholinergic neurons [7,8]. The other is an NMDA receptor antagonist, which inhibits brain cell death through excitotoxicity triggered by overstimulation of glutamate receptors [9]. Therefore, many considerable efforts are focused on the development and application of drugs as a novel therapeutic strategy for AD treatment.
In Korea, CKJ is predominantly made by short-term fermentation with Bacillus subtilis. CKJ contains various enzymes, microorganisms, and bioactive compounds that are absent from unfermented soybeans [10,11]. During fermentation, the hydrolysis reaction converts flavonoid glycosides into aglycones, and many proteins are broken down into small peptides and amino acid monomers [12,13]. Especially, CKJ has diverse biological and pharmacological activities, including anti-obesity, anti-diabetic, and anti-inflammatory effects on human chronic diseases. Significant reduction of blood glucose and glycosylated hemoglobin levels, as well as improved insulin tolerance, have been observed in CKJ-supplemented C57BL/Ksj-db/db mice, a type 2 diabetic animal model [14,15]. Further, reduction of passive cutaneous anaphylaxis was observed in CKJ-treated rat models of type I hypersensitivity and arachidonic acid-induced ear edema [16]. Furthermore, ethanol extracts of CKJ were shown to significantly increase the viabilities of cultured mice spleen and thymus cells by suppressing apoptotic cell death [17]. Although CKJ shows a variety of therapeutic activities in human chronic diseases, it has never been considered as a novel therapeutic strategy for AD treatment based on its ability to increase NGF metabolism.
Therefore, in this study, we investigated whether or not oral administration of CKJ significantly improves the AD phenotype in neuroblastoma cells as well as in the brain of AD mice having early stage. The data presented here provide strong evidence that CKJ is a potential candidate for the stimulation or enhancement of neuronal cell differentiation through activation of NGF biosynthesis and secretion in the brain.
Materials and Methods
Preparation of CKJ sample
Six different strains of soybeans (Pungsan; PS, Pungwon; PW, Daewon; DW, Daepung; DP, Taegwang; TG, and Shinhwa; SH) to produce CKJ sample were supplied by the National Institute of Crop Science in Miryang, Korea. B. subtilis used for the fermentation of CKJ was obtained from Applied and Environmental Microbiology Laboratory at Pusan National University. Whole soybeans (100 g) were washed and soaked with three volumes of tap water at room temperature for 12 h, followed by steaming for 30 min at 121℃. Thereafter, soybeans were allowed to cool to 37℃. The steamed soybeans were then inoculated with 5% (w/w) B. subtilis (1×109 cells/mL) and fermented for 48 h at 37℃. CKJ samples were freeze-dried and then homogenized, after which CKJ powder was stored at -75℃ before use.
Cell culture
The B35 neuronal cell line, which can secrete NGF, and the PC12 phenochromocytoma cell line were obtained from the Korean Cell Line Bank (Seoul, Korea). B35 cells were maintained for 24 to 36 h in Dulbecco's Modified Eagle Medium (DMEM, Hyclone, Logan, UT, USA) containing 10% fetal bovine serum (FBS, Hyclone), 100 IU/mL of penicillin, and 100 µg/mL of streptomycin. PC12 cells were cultured in RPMI 1640 (Hyclone) supplemented with 10% FBS, 100 IU/mL of penicillin, and 100 µg/mL of streptomycin. All cells were maintained in a humidified incubator at 37℃ and 5% CO2.
MTT assay
B35 cells were seeded at a density of 4×104 cells/200 µL in 96-well plates and grown for 24 h in a 37℃ incubator. When cells reached 70-80% confluence, they were exposed to distilled water (vehicle) or various types of CKJ extracts dissolved in distilled water for a further 24 h. Cell proliferation was determined using the tetrazolium compound MTT (3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide, Sigma-Aldrich, St. Louis, MO, USA). After discarding the supernatants in the vehicle or extract-treated wells, 200 µL of fresh DMEM and 50 µL of MTT solution (2 mg/mL in phosphate-buffered saline; PBS) were added to each well, after which cells were incubated in a 37℃ incubator. Reduction of MTT to insoluble purple formazan dye crystals by viable cells was evaluated in a 220 µL sample recovered after 4 h. The formazan precipitate was dissolved in DMSO, and the absorbance in the wells was read directly at 570 nm using a Soft Max Pro5 spectrophotometer (Molecular Devices, Sunnyvale, CA, USA). Data were analyzed in terms of cell number versus absorbance, thus allowing changes in cell proliferation to be quantified.
NGF detection using ELISA
To detect the levels of NGF in the culture supernatant and serum, the culture supernatant of B35 cells, which had been cultured and treated under the same conditions used in the MTT assay, as well as the serum from blood of Tg2576 mice were collected. The levels of NGF in the culture supernatant of B35 cells and serum were measured by using an ultra-sensitive assay and reagents in the rat NGF ELISA kit (Chemicon International Inc., Temecula, CA, USA). Briefly, the sample and standards were incubated overnight on antibody-coated plates in a shaker at 100-150 rpm at 2-8℃. The wells were then washed three times with wash buffer, after which 100 µL of anti-mouse NGF monoclonal antibody was added to each of the wells. The plates were then incubated in the shaker for 2 h at room temperature. The next step involved addition of 100 µL of peroxidase conjugated donkey anti-mouse IgG polyclonal antibody to each well and incubation at room temperature for 2 h. After washing, 100 µL of TMB/E substrate was added to each well, followed by incubation at room temperature for 15 min. The reaction was quenched by the addition of 100 mL of stop solution. The plates were analyzed by evaluating the absorbance at 450 nm using the same spectrophotometer.
Neuritic outgrowth assay
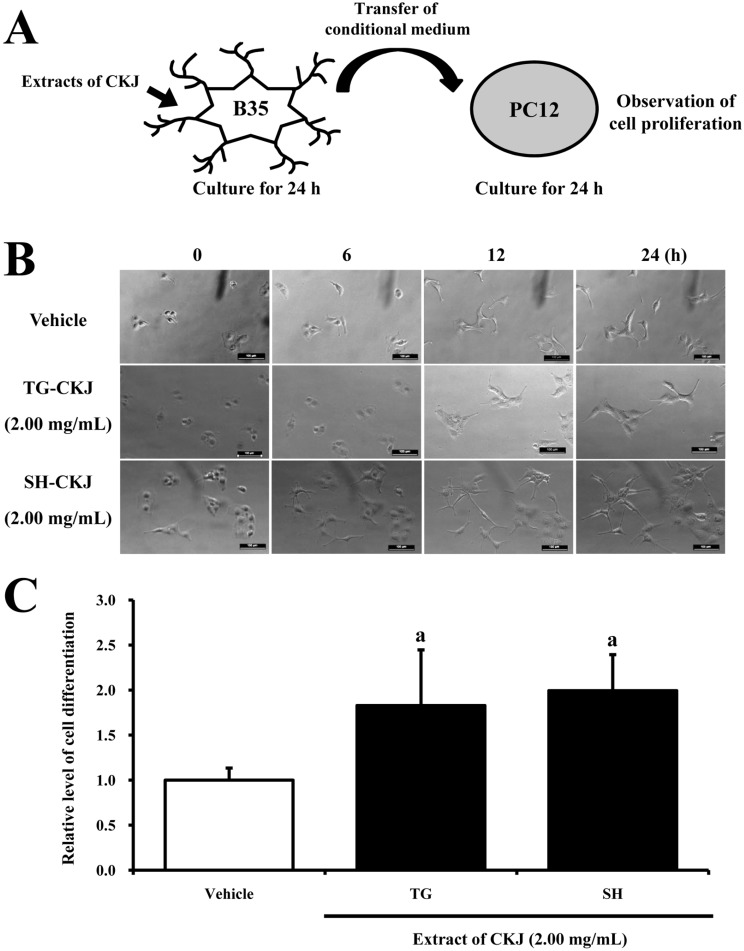
To confirm the effect of NGF on cell differentiation, CM was collected from B35 cells treated with TG-CKJ or SH-CKJ extract (2.00mg/mL), processed under a range of conditions for 24 h, and added to undifferentiated PC12 cells. After 24 h, the morphology of PC12 cells was observed by optical microscopy, and their dendrite length was measured using a Leica Application Suite (Leica Mycrosystems, Bannockbrun, IL, USA).
Western blot analysis
In the case of cells, PC12 cells were harvested from 100 mm-diameter culture dishes after being treated with TG-CKJ or SH-CKJ CM for 24 h. These cell pellets were solubilized in PRO-PREP protein extraction solution (INtRON Biotechnology, Seongnam, Korea) containing 1.0 mM phenylmethylsulfonyl fluoride (PMSF), 1.0 mM ethylenediamine tetraacetic acid (EDTA), 1 µM Pepstatin A, 1 µM Leupeptin, and 1 µM Aprotinin. In the case of brain tissue, frozen tissue was chopped with surgical scissors and homogenized with a glass homogenizer in PRO-PREP protein extraction solution. The resulting mixture was then centrifuged for 10 min at 13,000 rpm at 4℃. The homogenized proteins were separated for 2 h at 100 V by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to nitrocellulose membranes for over 2 h at 40 V. The membranes were then incubated with the following primary antibodies in order to determine the levels of each protein: anti-TrkA antibody (Cell Signaling Technology, Beverley, MA, USA), anti-p-TrkA antibody (Cell Signaling Technology), anti-Akt antibody (Cell Signaling Technology), anti-p-Akt antibody (Cell Signaling Technology), anti-Erk antibody (Santa Cruz Biotechnology, SantaCruz, CA, USA), anti-p-Erk antibody (Santa Cruz Biotechnology), anti-p75NTR antibody (Cell Signaling Technology), anti-RhoA antibody (Cell Signaling Technology), anti-Bcl2 antibody (Abcam, Cambridge, MA, UK ), anti-Bax antibody (Abcam), and anti-beta actin antibody (Sigma-Aldrich). Each membrane was washed with buffer (137 mM NaCl, 2.7 mM KCl, 10 mM NaHPO4, and 0.05% Tween-20) and incubated with a 1:1,000 dilution of horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG antibody at room temperature for 2 h. The membrane blots were developed using an Enhanced Chemiluminescence Reagent Plus kit (Amersham Life Science, Piscatway, NJ, USA).
Care and use of animals
The animal protocol used in this study was reviewed and approved based on the ethical and scientific care procedures of the Pusan National University-Institutional Animal Care and Use Committee (PNU-IACUC; Approval Number PNU-2012-0071). Female Non-Tg and Tg2576 mice on the B6SJLF/1 bacgkround were purchased from SamTacho (Osan, Korea) and handled at the Pusan National University Laboratory Animal Resources Center, which is accredited by the Korea FDA according to National Institutes of Health guidelines. All mice were given a standard irradiated chow diet (Purina Mills, Seoungnam, Korea) ad libitum and maintained in a specific pathogen-free state under a strict light cycle (lights on at 08:00 h and off at 20:00 h) at a temperature of 22±2℃ and at 50% relative humidity.
Experimental design
Fifteen-week-old Tg2576 mice (n=12) were assigned to one of two groups (n=6 per group): a vehicle treated group or SH-CKJ treated group. Vehicle treated Tg2576 mice received a constant volume of water per day via gavage, whereas SH-CKJ treated Tg2576 mice received 50 mg/kg of body weight/day of SH-CKJ extract diluted in distilled water via gavage for 8 weeks. Non-Tg mice without any treatment were used as a control group. After the final administration, animals were sacrificed immediately using CO2 gas to acquire blood and brain tissue. The samples were stored in Eppendorf tubes at -70℃ until assayed.
Perfusion and immunohistochemcal analysis
Brain perfusion and immunohistochemical analyses were performed as previously described [18,19]. Briefly, mice were anaesthetised with Zoletil 50 (Virbac, Carros cedex, France) and transcardially perfused with 1x PBS, followed by buffered 4% formaldehyde to effectively remove blood and fix brain tissues. After perfusion, the brain of each mouse was isolated from the skull and fixed overnight in formaldehyde. Each brain was then dehydrated and embedded in paraffin. A series of brain sections (10 µm) were cut from paraffin-embedded tissue using a Leica microtome (Leica Microsystems). For immunohistochemical analysis, these sections were deparaffinized with xylene, rehydrated, and pretreated for 30 min at room temperature with PBS blocking buffer containing 10% goat serum. Next, the sections were incubated with mouse anti-NGF antibody (Abcam) at a dilution of 1:100 in PBS blocking buffer. The antigen-antibody complexes were visualized with biotinylated secondary antibody (goat anti-rabbit)-conjugated HRP streptavidin (Histostain-Plus Kit; Zymed, South San Francisco, CA, USA) at a dilution of 1:1,500 in PBS blocking buffer. NGF protein was detected using stable 3,3'-diaminobenzidine (DAB; Invitrogen, Carlsbad, CA, USA) and observed using a model BX50F-3 optical microscope (Olympus, Tokyo, Japan).
Statistical analysis
One-way ANOVA was used to determine significant differences between the vehicle and CKJ treated groups (SPSS for Windows, Release 10.10, Standard Version, Chicago, IL, USA). Additionally, response differences between the Non-Tg and Tg2576 groups were evaluated by a post hoc test (SPSS for Windows, Release 10.10, Standard Version) of the variance and significance levels. All values are reported as the mean±SD. A p value of <0.05 was considered significant.
Results
Effects of CKJ manufactured by six different strains on B35 cell viability and NGF secretion ability
The effects of CKJ on viability and NGF secretion ability were evaluated in B35 cells treated with extracts of CKJ manufactured from six different soybean strains using MTT assay and NGF ELISA analysis. A significant alteration on the viability of B35 cells was not detected in any CKJ treated groups compared to the vehicle treated group, although some groups showed a slightly higher level on the cell viability (Figure 1A). However, the NGF concentration significantly increased in cells treated with TG-CKJ or SH-CKJ compared those treated with vehicle. On the other hand, those cells treated with other types of CKJ showed no change in NGF level (Figure 1B). These results suggest that only two types of TG-CKJ and SH-CKJ could induce elevation of NGF secretion ability along with no cytotoxicity in B35 cells. Therefore, these two types of CKJ were selected for further study, including dose-dependent effect assay, neuritic outgrowth assay, and NGF signaling pathway analysis.
Change along with the increase of TG-CKJ and SH-CKJ concentration on B35 cell viability and NGF secretion ability
In order to investigate an alteration along with the increase of TG-CKJ and SH-CKJ concentration, cell viability and NGF concentration were measured in B35 cells treated with four different concentrations of TG-CKJ or SH-CKJ. In the TG-CKJ treated group, cell viability was maintained within normal range in the 0.63, 1.25 and 2.50 mg/mL TG-CKJ treated group compared with the vehicle-treated group, whereas a significant decrease in cell viability was observed in only 5.00 mg/mL treated treated group (Figure 2A). Furthermore, the NGF concentration was significantly higher in only 2.50 and 5.00 mg/mL of the TG-CKJ treated groups compared to vehicle treatment (Figure 2B). In the SH-CKJ treated group, most concentrations of SH-CKJ except for 5.00 mg/mL resulted in maintained cell viability compared with vehicle treatment as TG-CKJ treated group (Figure 2C). In addition, maximum secretion of NGF was observed in the 1.25 mg/mL SH-CKJ treated group, followed by the 0.63, 2.50, and 5.00 mg/mL treated groups. But, a significant differences on the NGF secretion was detected in only 1.25 mg/mL SH-CKJ treated group (Figure 2D). These results indicate that TG-CKJ and SH-CKJ in some ranges of treated concentration may induce NGF secretion from B35 cells. Also, the commonly applicable concentrations of TG-CKJ and SH-CKJ for optimal secretion of NGF are considered to be 2.00 mg/mL which is a medium concentration between the optimal concentration of TG-CKJ and the optimal concentration of SH-CKJ.
Effect of CKJ on differentiation of PC12 cells
To determine whether or not NGF secreted from B35 cells in response to TG-CKJ or SH-CKJ treatment can induce differentiation of PC12 cells, the length of PC12 cells was observed after addition of CM collected from B35 cells treated with TG-CKJ or SH-CKJ. Differentiation of PC12 cells was enhanced with time in all groups, including vehicle, TG-CKJ, and SH-CKJ treated groups. However, the length of PC12 cells at 24 h was significantly higher in the TG-CKJ and SH-CKJ CM treated groups compared to the vehicle treated group (Figure 3). Therefore, the above results suggest that NGF secreted from B35 cells treated with TG-CKJ or SH-CKJ can induce differentiation of PC12 cells.
Effect of CKJ treatment on NGF receptor TrkA signaling pathway in PC12 cells
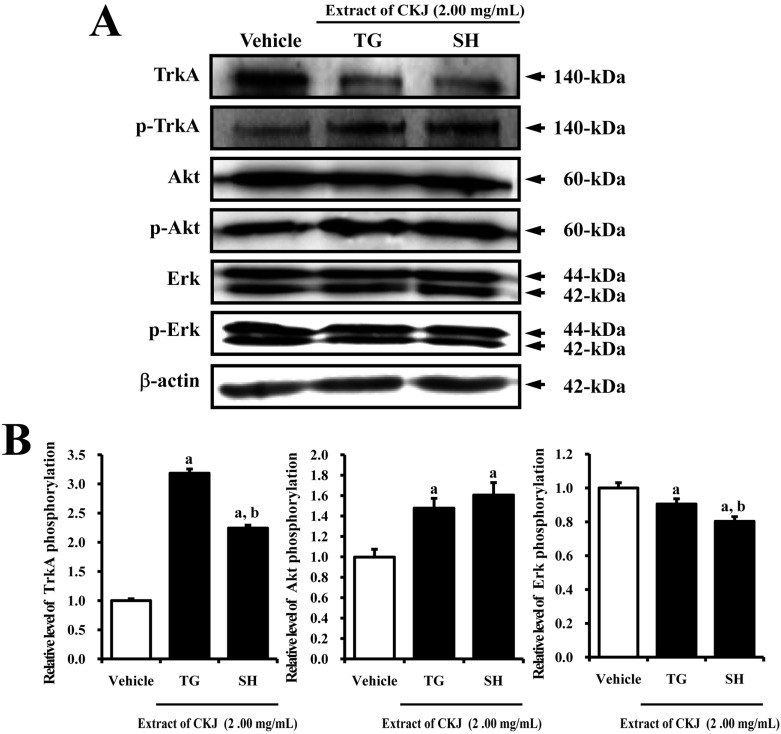
The NGF secreted from neuronal cells transfers its activation signal into the cytosol by binding to two types of NGF receptors located on the cell membrane. Of the two types of NGF receptors, the TrkA high affinity receptor can induce neuritic outgrowth via the Akt and Erk signaling pathways [20]. Therefore, to examine whether or not NGF secreted from B35 cells following TG-CKJ and SH-CKJ treatment can induce activation of the TrkA receptor signaling pathway, the expression levels of several protein were measured in PC12 cells cultured with CM. The phosphorylation level of TrkA was higher in the TG-CKJ and SH-CKJ CM treated groups compared to the vehicle CM treated group, although the rate of increase was greater upon TG-CKJ CM treatment compared to SH-CKJ CM. Upon activation, TrkA may transfer its signal via two different pathways, the Akt- and Erk-mediated pathways, for induction of cell differentiation and survival. Alteration of Akt phosphorylation was very similar with that of TrkA phosphorylation. Especially, the SH-CKJ CM treated group showed greater Akt phosphorylation than the TG-CKJ CM treated group. However, Erk phosphorylation was slightly decreased in the TG-CKJ and SH-CKJ CM treated groups compared with the vehicle treated group (Figure 4). These results suggest that NGF in TG-CKJ or SH-CKJ CM can induce differenciation of PC12 cells through stimulation of TrkA and Akt phosphorylation in the high affinity NGF receptor TrkA signaling pathway.
Effect of CKJ treatment on NGF receptor p75NTR signaling pathway in PC12 cells
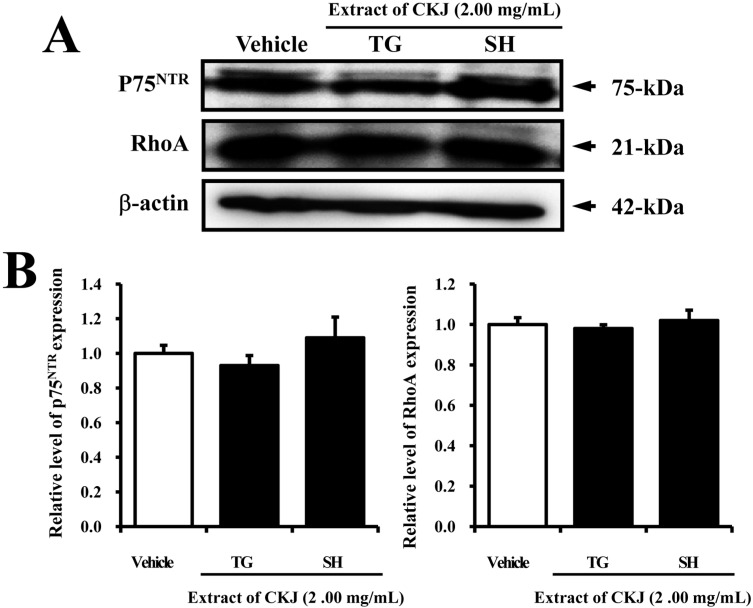
The effects of NGF secreted into TG-CKJ and SH-CKJ CM on the low affinity NGF receptor p75NTR signaling pathway were examined in PC12 cells. There was no significant difference between any CKJ CM treated groups and the vehicle treated group in the expresson of NGF receptor p75NTR, although the TG-CKJ CM treated group showed slightly lower expression compared to the SH-CKJ CM treated group. The overall pattern of RhoA expression was very similar with that of p75NTR expression (Figure 5). These results show that NGF in TG-CKJ and SH-CKJ CM may not induce any significant change in the NGF receptor p75NTR signaling pathway. Based on all in vitro results, we selected SH-CKJ in order to investigate the effect of CKJ on Tg2576 mice showing an early stage of AD phenotypes.
Effect of SH-CKJ treatment on NGF secretion ability in the brain of Tg2576 mice
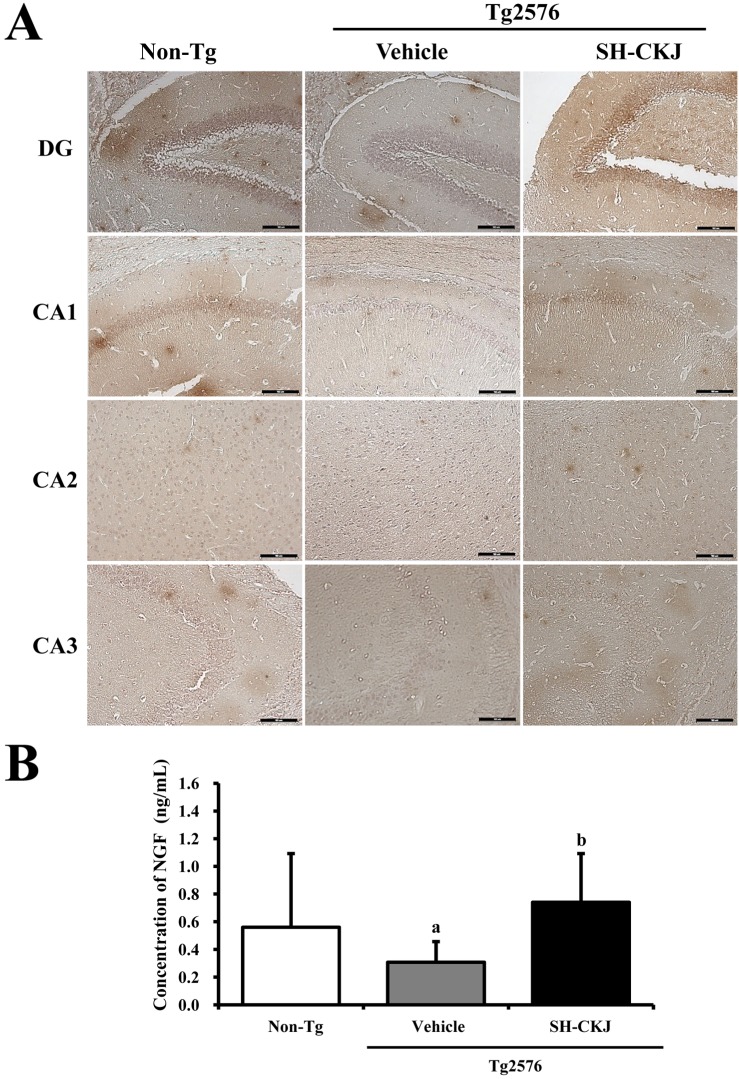
To assess whether or not SH-CKJ treatment for 8 weeks could induce an increase in NGF secretion, NGF levels were measured in the serum and brain tissue of Tg2576 mice using ELISA analysis and immunostaining. NGF antibody appeared to be lowest reactive in the vehicle treated Tg2576 mice, with high densities in CA1, CA2, and dentate gyrus (DG), compared with Non-Tg groups. After SH-CKJ treatment, the density of NGF-stained cells was significantly elevated in CA1, CA2, and DG of SH-CKJ treated Tg2576 mice, whereas the CA3 region maintained a constant level (Figure 6A). Furthermore, alteration of the serum level of NGF showed an identical pattern as that observed in the immunostaining analysis. There was a significant decrease in the serum level of NGF in vehicle treated Tg2576 mice compared with Non-Tg mice. In SH-CKJ treated Tg2576 mice, the serum level of NGF was recovered to that of Non-Tg mice (Figure 6B). Therefore, the above results indicate that SH-CKJ treatment may induce stimulation of NGF production in the serum and brain tissue of Tg2576 mice.
Effect of SH-CKJ treatment on NGF receptor TrkA signaling pathway in Tg2576 mice
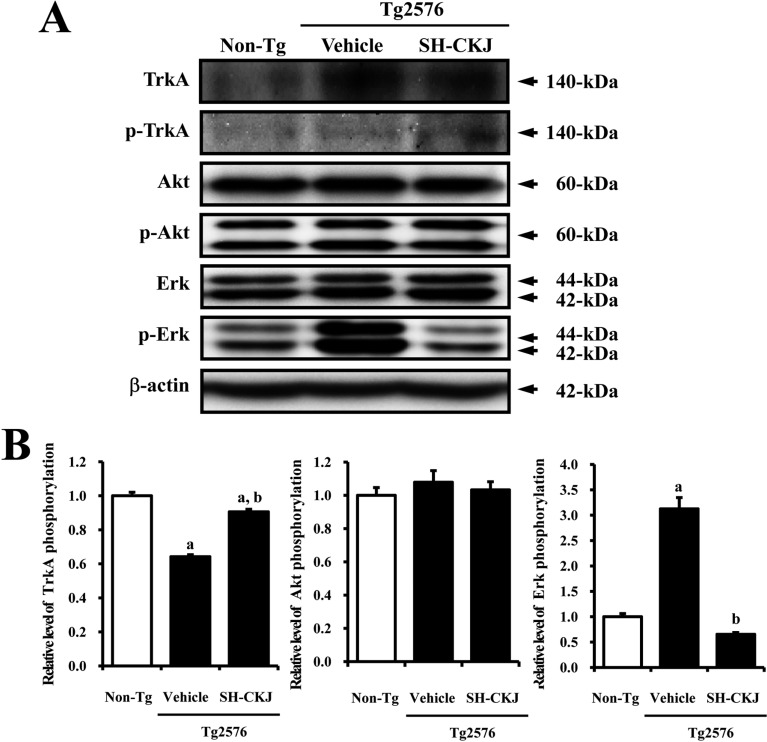
To investigate the effect of SH-CKJ treatment on the NGF receptor TrkA signaling pathway, the expression levels of several proteins were measured in brain tissue of Tg2576 mice. A dramatic change in the phosphorylation levels of TrkA and Erk similar to those in vitro was detected, whereas the phosphorylation level of Akt was maintained. The phosphorylation level of TrkA was lower in vehicle treated Tg2576 mice compared to Non-Tg mice, whereas that of Erk showed the reverse pattern. However, in SH-CKJ treated Tg2576 mice, these levels were dramatically recovered to those observed in Non-Tg mice. Especially, the phosphorylation level of Erk was elevated by 312% in Tg2576 mice, whereas that of TrkA was reduced by 36%. On the other hand, the phosphorylation level of Akt was maintained without any significant change (Figure 7). These results suggest that SH-CKJ treatment may lead to recovery of TrkA and Erk phosphorylation in the high affinity NGF receptor TrkA signaling pathway in brain tissue of Tg2576 mice.
Effect of SH-CKJ treatment on NGF receptor p75NTR signaling pathway in Tg2576 mice
Finally, to investigate whether or not NGF treatment can affect the NGF receptor p75NTR signaling pathway, the expression levels of key components were examined in brain tissue of Tg2576 mice after treatment for 8 weeks. Surprisingly, significant alteration of the expression levels of four components, including p75NTR, RhoA, Bcl-2 and Bax, was not detected in any of the groups (Figure 8). Therefore, these results show that SH-CKJ treatment may not induce any significant change in the NGF receptor p75NTR signaling pathway in Tg2576 mice.
Discussion
NGF is a member of the neurotrophic family of factors and carries out an important role in the growth, survival, and maintenance of nerve cells [21]. Especially, it plays a vital role in axonal growth and the survival of sympathetic and sensory neurons [22,23]. Furthermore, NGF has powerful beneficial effects on damaged or dying neurons in neurodegenerative diseases such as AD and dementia, as elevating the levels of appropriate neurotrophic factors can help to restore the health and vitality of injured neurons [24]. However, NGF has not been applied for medical applications until now since this molecule cannot penetrate the blood-brain barrier owing to its large molecular weight and susceptibility. Therefore, most studies have focused on the development of novel agents that have the ability to increase the level of NGF secretion or enhance NGF activity. In this study, we investigated the effects of CKJ, a commonly consumed fermented food in Korea, on NGF secretion from neurobalstoma cells as well as transgenic mice maintaining a low level of NGF. Our results showed that secretion of NGF and its metabolism were enhanced by some types of CKJ, although the increase ratio varied among the groups.
Meanwhile, enhancement of NGF secretion and neuritic outgrowth of PC12 cells can be induced by extracts of certain medicinal herbs, including Liriope platyphylla and Cistanches Herba (CHE) [25,26]. Especially, Uezato et al. [27] screened a total of 120 samples to identify the effects of natural medicines on neuritic outgrowth in PC12 cells. Five natural medicines, including Trichosanthes root, Asiasarum root, Lycium bark, Sinomenium stem, and Dictamni radices, significantly promoted not only neuritic outgrowth but also stabilized neuritic formation in PC12 cells. Furthermore, various compounds isolated from soybeans were shown to be tightly associated with the secretion of neurotrophic factors and neuritic outgrowth. The administration of estradiol and soy phytoestrogens has been found to induce increase BDNF mRNA expression in the frontal cortex of female rats compared to ovariectomized controls [28]. Genistein, an isoflavone, induced NGF-induced neuritic outgrowth in PC12 cells through activation of Na+/K+/2Cl- cotransporter isoform 1 (NKCC1), although there was little effect on NKCC1 expression [29]. In addition, the effect of the flavonoid quercetin on neuritic outgrowth was investigated in PC12 cells. The above results suggested that quercetin stimulates NGF-induced neuritic outgrowth through enhancement of Cl(-) incorporation in the intracellular space using activated NKCC1 [30]. However, little is known about the use of CKJ for treatment of neurodegenerative diseases as well as their direct effects on NGF secretion both in vitro and in vivo. The results of our study are in agreement with the above reports, although the treatment materials and rates of increase varied. Further, CKJ used in this study contained various compounds originated from unfermented soybeans and synthesized by fermentation (data not shown), whereas previous studies used only single compounds.
NGF secreted from neuronal cells has been shown to induce differentiation and growth of cells through the NGF receptors TrkA and p75NTR [31,32]. CKJ-induced NGF and CHE-induced NGF are similar in terms of their activation of the NGF receptor signaling pathway. CHE extract was previously shown to induce an increase in NGF secretion in C6 cells, whereas NGF bound to TrkA receptor and stimulated neuritic extension in PC12 cells [26]. In our study, CKJ-induced NGF stimulated neuritic outgrowth and differentiation of PC12 cells through NGF receptor TrkA rather than p75NTR. Furthermore, the same results were observed in brain tissue of Tg2576 mice.
There are very few studies on NGF secretion ability in animals. Spicatoside A isolated from Liriope platyphylla effectively induced enhancement of NGF in the serum of an animal model [33]. Furthermore, CHE extracts have been shown to induce neuronal cell differentiation, increased neuritie length, and synapse formation in the hippocampus of mice as well as enhancement of learing and memory [26]. In this study, we used Tg2576 mice expressing an AD phenotype, including behavioral defects, increased Aβ-42 production, and formation of amyloid plaques, to investigate the effects of CKJ. Our results are the first ever to demonstrate the possibility that SH-CKJ can improve AD symptoms through enhancement of NGF secretion, although further study is needed to verify the detailed mechanism.
Tg2576 mice overexpressing the Swedish mutant form of human APP695 show AD-like phenotypes, including behavioral defects, a 14-fold increase in Aβ-42 peptide levels, and formation of neumerous amyloid plaques at 9 to 10 months of age [34]. Further, the level of NGF mRNA was shown to be decreased by 44% in 10-month-old Tg2576 mice, whereas the level of p75NTR mRNA was reduced by 25% [35]. Our results are completely in agreement with previous studies despite our detection method being different. However, Chauhan and Siegel did not detect the protein level of NGF receptor TrkA, a key component in the signaling pathway [35]. Our study is the first to detect two types of NGF receptors and their signaling pathways at the protein level. Therefore, the current study provides new evidence that CKJ could induce the expression of TrkA receptor and its signaling pathway.
Take together, these results suggest that treatment with CKJ containing enhanced flavonoid and phenolic compounds for 8 weeks can stimulate NGF secretion and the NGF receptor TrkA signaling pathway in PC12 cells and Tg2576 mice. Therefore, CKJ may be considered as a benefit extract to the improvement of NGF metabolism.



 XML Download
XML Download