

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Ginkgo biloba extract (EGb 761) is derived from the green leaves of Ginkgo biloba and used as a traditional medicine for various purposes [1]. It consists of flavones glycosides and terpene lactones and has an antioxidant effect in ischemic brain injury [2,3]. EGb 761 also provides pharmacologic benefits in Alzheimer's disease and vascular disorders of the brain [4-6], acts as a scavenger of reactive oxygen species (ROS), and reduces neural cell damage induced by ischemia [7,8]. Moreover, previous studies clearly reported the neuroprotective effect of EGb 761 during focal cerebral ischemia induced by middle cerebral artery occlusion (MCAO) [9,10].
Parvalbumin belongs to the family of EF-hand Ca2+-binding proteins, and acts as a calcium sensor protein or calcium buffer protein because its critical function is binding to Ca2+ molecules released into the cytoplasm [8,11,12]. Thus, parvalbumin plays an essential role in maintaining calcium homeostasis of the central nervous system by decreasing the intracellular Ca2+ concentration [12]. An increase in intracellular Ca2+ concentration leads to neuronal cell death and consequently central nervous system disorder [13]. Parvalbumin is thought to exert a neuroprotective effect by preventing increases in intracellular Ca2+. Although previous studies have attempted to elucidate the physiologic mechanism underlying the neuroprotective effect of EGb 761, more information is needed in order to fully define the complete neuroprotective mechanism. We hypothesized that EGb 761 may mediate parvalbumin expression in ischemic brain injury. However, little data is available regarding the effects of EGb 761 on parvalbumin expression levels in neuronal cell injury. We investigated the effect of EGb 761 on parvalbumin during cerebral ischemic injury.
Materials and Methods
Animals and EGb 761 treatment
All animal experiments were followed a protocol approved by the Institutional Animal Care and Use Committee (IACUC) at the Gyeongsang National University. Male Sprague-Dawley rats (210-230 g, n=48) were purchased from Samtako Co. (Animal Breeding Center, Osan, Korea). Animals were maintained on 12 h light/dark cycle with free access to food and water. Animals were randomly divided into following three groups: sham-operated group, vehicle-treated group, and EGb 761-treated group (n=16 per group). Animals were intraperitoneally administrated EGb 761 (100 mg/kg, Yuyu, Seoul, Korea) in EGb 761-treated group and vehicle in the vehicle-treated group at 1 h before the onset of MCAO [9].
Experimental model of cerebral ischemic injury
Focal cerebral ischemia was induced by the method of MCAO [14]. Before MCAO operation, animals were anesthetized with sodium pentobarbital (30 mg/kg). a midline incision was made and The right common carotid, external carotid artery and internal carotid arteries were exposed. The external carotid artery was tied and cut. A 4/0 monofilament nylon with its tip rounded by heating over a flame was advanced from the external carotid artery into lumen of the internal carotid artery, thereby occluding blood flow through the middle cerebral artery. At 24 h after the onset of permanent occlusion, brain tissues were removed.
Triphenyltetrazolium chloride (TTC) staining
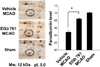
Brains were cut into coronal slices of 2 mm in thickness. These slices were stained for 20 min in a 2% triphenyltetrazolium chloride (TTC; Sigma, St. Louis, MO, USA) and fixed in 10% formalin. The stained slices were photographed by a Nikon CoolPIX990 digital camera (Nikon, Tokyo, Japan) and measured for the ischemic lesion by Image-ProPlus 4.0 software (Media Cybernetics, Silver Spring, MD, USA). The ischemic lesion percentage of each slice was calculated by the ratio of the infarction area to the whole slice area.
Two-dimensional gel electrophoresis
The right cerebral cortices were dissolved in lysis buffer (8 M urea, 4% CHAPS, ampholytes, and 40 mM Tris-HCl). After centrifuge at 16,000 g for 20 min at 4℃, the total protein concentration was determined using the Bradford method (Bio-Rad, Hercules, CA, USA) according to the manufacturer's protocol. Briefly, prior to isoelectric focusing (IEF), the immobilized pH gradients (IPG, pH 4-7, pH 6-9, 17 cm, Bio-Rad) gel strips were re-hydrated in buffer (8 M urea, 2% CHAPS, 20 mM DTT, 0.5% IPG buffer, bromophenol blue) for 13 h. The protein samples (100 µg) were loaded on IPG strips. IEF was followed a multi-step protocol: 250 V (15 min), 10000 V (3 h), and then 10000 V to 50000 V using Protean IEF Cell (Bio-Rad). After equilibration of the isoelectric focusing strips, IPG strips were applied onto gradient gels (7.5~17.5%) and electrophoresed at 10 mA for 10 h using Protein-II XI electrophoresis equipment (Bio-Rad) at 10℃. The gels were fixed in a solution (12% acetic acid, 50% methanol) for 2 h, washed twice with 50% ethanol for 20 min, and then sensitized in 0.2% sodium thiosulfate for 1 min. The gels were washed with deionized water, incubated in a silver solution (0.2% silver nitrate, 0.75 mL/L formaldehyde) for 20 min. The gel was developed in a solution (0.2% sodium carbonate, 0.5 mL/L formaldehyde) and the reaction was stopped by adding 1% acetic acid. The silver stained gels were scanned by Agfar ARCUS 1200™ (Agfar-Gevaert, Mortsel, Belgium) and analyzed by a standard protocol for PDQuest software (Bio-Rad). The analyzed data were expressed as mean±S.E.M. Relative intensity of spots in each group was analyzed and compared by Student's t-test. Alteration of protein expression was identified in vehiclel- and EGb 761-treated animals. The gel pieces containing the desired protein spots were manually cut out from the gels and processed for MALDI-TOF.ed and distained. Detained and dehydrated gel pieces were incubated with trypsin-containing digestion buffer for 24 h at 37℃. The peptides were extracted and analyzed in a Voyager-DE™ STR biospectrometry workstation (Applied Biosystem, Forster city, CA, USA) for MALDI-TOF mass spectrometric analysis. the acquired peaks were identified using search programs MS-Fit and ProFound. SWISS-PROT and NCBI were used as the protein sequence databases.
RNA isolation and reverse transcription-PCR amplification
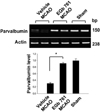
RNA was extracted using Trizol reagent according to the manufacturer's instruction (Invitrogen, Carlsbad, CA, USA). Total RNA (1 µg) was reverse transcribed using superscript III first-strand system for RT-PCR (Invitrogen) based on the manufacturer's protocol. The primers sequences of parvalbumin are 5'-AAGAGTGCGGATGATGTGAAG-3' (forward primer) and 5'-AGCCATCAGCGTCTTTGTTT-3' (reverse primer). The primers sequences of actin are 5'-GGGTCAGAAGGACTCCTACG-3' (forward primer) and 5'-GGTCTCAAACATGATCTGGG-3' (reverse primer). The PCR reaction was carried out as followed: 5 min at 94℃, 30 sec at 94℃, 30 sec at 54℃, 1 min at 72℃ and 10 min at 72℃. The samples were amplified 30 cycles. PCR product was run in a 1% agarose gel and visualized under UV light.
Protein preparation and Western blot analysis
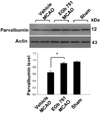
Western blot analysis were performed as previously described [10]. The proteins of right cerebral cortex were extracted by homogenization in buffer [1% Triton X-100, 1 mM EDTA in 1×PBS (pH 7.4)] containing 200 µM phenylmethylsulfonyl fluoride and 10 µM leupeptin. The lysates were centrifuged at 12,000 rpm for 20 min at 4℃. The supernatants were collected and the protein concentration of each lysate was determined using the bicinchoninic acid (BCA) kit (Pierce, Rockford, IL, USA). Total protein (30 µg) was run on to 10% SDS-polyacrylamide gel and subsequently transferred to a poly-vinylidene fluoride (PVDF) membranes (Millipore, Billerica, MA, USA) and membranes were blocked with 5% skim milk in 0.1% Tween-20 (TBST). After washing membranes with TBST, membranes were incubated with the following antibodies: a rabbit polyclonal anti-rat parvalbumin IgG (diluted 1:1000, Cell Signaling Technology, Beverly, MA, USA) and a mouse monoclonal anti-rat actin IgG (diluted 1:1000, Santa Cruz Biotechnology, Santa Cruz, CA, USA). The membrane was incubated with secondary antibody (1:5000, Pierce, Rockford, IL, USA) and the ECL Western blot analysis system (Amersham Pharmacia Biotech, Piscataway, NJ, USA) was used for detection according to the manufacturer's protocol.
Immunohistochemical staining
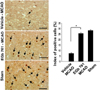
Brains were fixed in 4% paraformaldehyde in 0.1M phosphate-buffered saline (PBS, pH 7.4) and embedded in paraffin. Three sections were selected in each group for the immunohistochemical staining. Sections were blocked with 1% normal goat serum in PBS for 1 h and incubated with a rabbit polyclonal anti-rat parvalbumin IgG (diluted 1:200, Cell Signaling Technology, Beverly, MA, USA) at 4℃ for 15 h. After washing with PBS, sections were incubated with biotin-conjugated goat anti-rabbit IgG (1:200 in PBS) for 1 h, followed by incubation with an avidin-biotin-peroxidase complex for 1 h from a Vector ABC Elite kit (Vector Laboratories Inc., Burlingame, CA, USA), incubated with DAB solution with 0.03% hydrogen peroxidase. Sections were counterstained with hematoxylin, dehydrated in graded alcohol. For the positive cell counting, three fields for cortical region were selected from right cerebral cortex. The total cell number and parvalbumin-positive cell number were obtained in each field. The index of positive cells was described as the percentage of the number of parvalbumin-positive cells to the total number of cells in each field.
Statistical analysis
All data are expressed as mean±S.E.M. The intensity analysis of Western blot analysis was carried out using SigmaGel 1.0 (Jandel Scientific, San Rafael, CA, USA) and SigmaPlot 4.0 (SPSS Inc., Point Richmond, CA, USA). The results in each group were compared by one-way analysis of variance (ANOVA) followed by Student's t-test. The difference for comparison was considered significant at *P<0.05.
Results
We confirmed the neuroprotective effect of EGb 761 against ischemic brain injury using TTC staining (Figure 1). EGb 761 treatment significantly reduces infarct volume, compared with the vehicle-treated animals. Our previous study showed the EGb 761 regulates various proteins including peroxiredoxin-2, protein phosphatase 2A subunit B, and collapsing response mediator protein 2 in cerebral ischemic injury [15]. In this study, a proteomic approach additionally demonstrated that parvalbumin protein spots were differentially expressed in the cerebral cortices of vehicle- and EGb 761-treated animals in which brain injury had been induced by MCAO (Figure 2). The peptide mass of parvalbumin is 5/103 and the sequence of this protein is 40%. The intensity of parvalbumin protein spots were decreased in vehicle-treated animals, whereas this decrease was prevented in EGb-761-treated animals. The parvalbumin levels were 0.49±0.02 and 0.83±0.02 in the vehicle- and EGb 761-treated animals, respectively. RT-PCR and Western blot analyses clearly demonstrated differences in parvalbumin levels between the vehicle- and EGb 761-treated animals. Parvalbumin transcript levels were 0.28±0.03 and 0.87±0.02 in the vehicle- and EGb 761-treated animals during MCAO, respectively (Figure 3). Parvalbumin expression was significantly decreased in vehicle-treated animals, whereas EGb-761 treatment attenuated the ischemic injury-induced decrease in parvalbumin expression. Parvalbumin protein levels were 0.62±0.03 and 0.92±0.02 in the cerebral cortices of vehicle- and EGb 761-treated animals during MCAO, respectively (Figure 4). Immunohistochemical staining showed that the number of parvalbumin-positive cells was lower in vehicle-treated animals than in sham-operated animals, and EGb 761 prevented the injury-induced decrease in parvalbumin-positive cells (Figure 5). The number of parvalbumin positive cells significantly decreased in vehicle-treated animal, whereas EGb761 treatment attenuates the injury-induced decrease of parvalbumin positive cells. The index of positive cells were 7.58±1.05% and 25.36±1.35% in vehicle- and EGb 761-treated animals during MCAO (Figure 5).
Discussion
EGb 761 has a neuroprotective effect against beta-amyloid toxicity and oxidative stress [16,17]. Our previous study clearly showed the neuroprotective effect of EGb 761 against focal cerebral ischemia induced by MCAO [14]. EGb 761 exerts a neuroprotective effect in various neurological disorders such as ischemia, Alzheimer's, and depression [5,18,19]. Moreover, we demonstrated that EGb 761 prevents neuronal cells death from ischemic brain injury through the specific modulation of various proteins including peroxiredoxin-2, protein phosphatase 2A subunit B, collapsing response mediator protein 2, adenosylhomocyteinase, pyruvate kinase isoenzyme, and isocitrate dehydrogenase. This study additionally showed the decrease of parvalbumin expression during MCAO. Parvalbumin is a calcium buffering protein that counteracts neuronal cells damage and exerts a neuroprotective effect by buffering intracellular calcium [20]. The intracellular Ca2+ concentration increases in ischemic brain injury, and this increase in Ca2+ induces lethal effects such as free radical production, which ultimately lead to neuronal cell degeneration and death [13,21]. Because an excessive intracellular calcium concentration leads to the initiation of neuronal cell death, the regulation of calcium concentration is critical for ensuring the survival of neuronal cells [22,23].
The proteomic approach revealed in this study revealed that EGb 761 prevents the brain injury-induced decrease in parvalbumin expression during MCAO. The maintenance of parvalbumin levels may inhibit the excessive increase in the intracellular calcium concentration that is induced by ischemia. Calcium binding proteins play an important role in the destiny of neuronal cells because they act as calcium chelators and reduce intracellular calcium concentration [24,25]. The rise of intracellular calcium activates calcium-sensitive protein digesting enzymes and leads to cellular degradation. Moreover, amyloid-beta induces the disruption of neuronal Ca2+ homeostasis and the generation of ROS, while these events are blocked by EGb 761 [7]. Western blot analysis and immunohistochemical staining clearly confirmed that cerebral ischemic injury decreases parvalbumin expression, whereas EGb 761 attenuates this injury-induced decrease in parvalbumin expression. Our results demonstrate that EGb 761 modulates parvalbumin expression in ischemic brain injury and consequently contributes to the regulation of intracellular calcium levels. Although further research is required to explain the relationship between EGb 761 and parvalbumin, these results suggest that EGb 761 maintains parvalbumin levels, thereby maintaining intracellular calcium levels and attenuating neuronal cell death. In conclusion, our findings suggest that EGb 761 attenuates neuronal cell death by modulating the expression of parvalbumin.

 XML Download
XML Download