

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Estrogen receptors (ERs) belong to the steroid hormone superfamily of nuclear receptors (NRs) [1,2]. Other types of steroid receptors among the NRs include the estrogen-related receptors (EER), progesterone receptors (PR), androgen receptors (AR), glucocorticoid, and mineral corticoid receptors. Steroid receptors (SRs), act as ligand-dependent transcription factors, and their activity is associated with the cell cycle [3]. Analyses of the association between steroid receptor activity and the cell cycle revealed that SRs, especially ERs, regulate cell proliferation [4]. Moreover, estrogen is an important sex hormone produced primarily in the ovaries in females and testes in males. This steroid hormone regulates the growth, development, and physiology of the reproductive system in humans [5]. Estrogen also affects the neuroendocrine, skeletal, adipogenic, and cardiovascular systems [6,7]. The biological functions of estrogen are mediated by binding to the ERs: estrogen receptor alpha (ERα) and estrogen receptor beta (ERβ). Estrogen signaling is selectively stimulated or inhibited depending upon a balance between ERα and ERβ activities in target organs. ERα was cloned from human breast cancer MCF-7 cells in 1986 and ERβ was cloned from rat prostate in 1996 [8].
ERα and ERβ are encoded by distinct genes located on different chromosomes. The human ERα gene is located on chromosome 6 while the ERβ gene is on chromosome 14 [9]. The full-length human ERα protein has 595 amino acids and a molecular size of 66 kDa while the full-length human ERβ protein has 530 amino acids and a molecular size of 54 kDa. Similar to other NRs, ERs have five domains with distinct functions [10]. The N-terminal of the A/B domains of ERs consist of activation function 1 (AF1), which contributes to the transcriptional activity of ERs and is an essential domain for interaction with co-regulators. AF1 is the least conserved region with only 30% identity between ERα and ERβ. Functional studies have shown that ERβ has low levels of AF1 activity. The A/B domains also contain amino acids that are targets of post-transcriptional modifications including splicing to stimulate AF1 activity [9]. The C domain encodes a centrally located DNA binding domain (DBD) essential for sequence-specific binging of ERs to DNA and regulating the expression of target genes [11]. The D domain, a hinge region, includes amino acid sequences that stimulate nuclear localization signaling and facilitate post-translational modification of ERs, resulting in the activation of ER signaling in cells. Finally, the E/F domain, located in the C-terminal region, contains a ligand-binding domain (LBD) that serves as an interaction site with co-regulators and ligand-dependent activation function 2 (AF2). AF1 and AF2 control the transcriptional regulatory activity of ERs because activation of ERs are stimulated during cellular responses to the environment [12]. The E/F domains of ERα and ERβ share a 53% sequence identity and affect cellular responses through ligand-dependent ER activation. The F domain also affects the activity of ERα and ERβ. The differences between the F domain of the ERs may contribute to the ability of ERs to selectively control transcriptional activities of specific target genes [10].
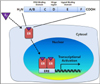
In the absence of hormones, ERs remain in an inactivation form due to association with heat shock protein 90 (Hsp90). Hsp90 regulates various processes in eukaryotic cells including protein stabilization, binding affinity of receptors to ligands, and signaling cascades [13]. Hsp90 inhibits the degradation of unbound ERs and prevents inactivated ERs from binding ligand [14]. After binding to ligand, ERs are phosphorylated, form homo- or heterodimers, and then translocate into the nucleus [15]. ERs modulate the transcription of target genes by binding to estrogen response elements (EREs) in the DNA sequence [16]. Binding of ERs to EREs promotes DNA bending and looping, thereby permitting interaction with the transcriptional machinery and co-regulator proteins. These co-regulators include co-activators, co-repressors, co-integrators, histone acetyltransferases and deacetylases, and general transcriptional factors [12]. ER/co-regulator complexes act specifically on target genes in particular organs according to extracellular stimuli [17]. Analysis of ERα and ERβ tissue distribution suggests that ERs have high specificity on the target tissue [18]. ERα is highly expressed in the uterus, prostate stroma, ovarian theca cells, Leydig cells in testes, epididymis, breast, and liver [19]. ERβ is highly expressed in prostate epithelium, testes, ovarian granulosa cells, bone marrow, and brain [20]. As mentioned above, ERα and ERβ have different downstream transcriptional activities, resulting in their tissue-specific biological actions [21].
The reproductive system in ER knockout mice
ERα and ERβ have different biological functions. Not surprisingly, eliminating the expression of either factor results in distinct phenotypes in mice. Analysis of ERα and ERβ using knockout mouse models has demonstrated the general roles of estrogen signaling. ERα (αERKO) and ERβ (βERKO) knockout mice have been used to examine the various roles of ER signaling [22,23].
Fertility
Rodent reproductive physiology is associated with steroid hormones, including estrogen. Estrogen is basically known for its effect on the reproduction systems of both females and males [24]. Understanding of the general physiological roles of estrogen has resulted in ovariectomies to ablate estrogen production or restoring estrogen levels with exogenous E2 [25]. Currently, the exact function of estrogen signaling in biological processes is unclear. However, transgenic mice have been useful for discovering gene-specific ER functions [26]. Many experiments with ERKO mice were able to elucidate ERs function in specific pathological conditions, absence of ER expression. [27,28]. Surprisingly, both male and female αERKO mice are infertile, whereas fertility differs among βERKO mice according to gender. Female βERKO mice have reduced fertility while males have normal fertility [29].
Reproductive organs
Generally, maturation of the reproductive organs promoted by E2 is important for successful pregnancy and appropriate sexual behavior [30]. Infertility in αERKO female mice is due to a failure to respond to estrogen in the uterus, which is a central organ for reproduction and pregnancy [31]. ERα is essential for uterine maturation but not development [32]. Female αERKO mice have a decreased number of glands in the endometrium, whereas female βERKO mice have normal uterine and vaginal tissues despite their reduced fertility rates. Another reason for infertility in the knockout mice is abnormal ovarian function. The ovary is affected by changes in estrogen levels that induce cyclic ovulation [33]. Estrogen stimulates follicular responses to gonadtropins and activities of enzymes such as aromatase for steroidogenesis [34]. The synthesis and secretion to E2 is known for action of follicular stimulating hormone (FSH) on granulosa cells and luteinizing hormone (LH) on theca cell. Both FSH and LH maintain the menstrual cycle and affect ovulation [35]. An abnormal ovarian phenotype causes infertility in αERKO mice and reduced fertility in βERKO mice. αERKO mice appear incapable of ovulation as their follicles fail to differentiate. Ovarian dysregulation is also observed in βERKO mice including an increased number of unruptured follicles and reduced numbers of oocytes [36].
Mammary gland
In mammals, the mammary gland is important for feeding offspring through the production milk. Development of the mammary gland involves ductal elongation from a nipple and formation of branches of the glands with fat pads. During the estrous cycle, mammary gland development proceeds in a manner similar to that observed during pregnancy through the actions of ERs and PRs [37]. Since αERKO female mice cannot ovulate, mammary glands of these mice do not develop beyond an immature state, and only contain rudimentary ducts characteristic of embryonic and fetal stages. Insufficient maturation of the mammary gland is caused by a loss of estrogen signaling due to the absence of ER expression. In female αERKO mice, the release of prolactin from the anterior pituitary gland is reduced and breast tissue for lactation is structurally impaired. However, the βERKO female have ability to nurse offspring since they develop normal mammary glands capable of lactation [38].
Male reproduction
The balance of two steroid hormones, androgen and estrogen, plays an important role in male reproductive organs, and is essential for normal prostate development and prostatic homeostasis [39]. However, estrogen signaling also plays a role in maintaining the male reproductive system. ERα is expressed in prostatic tissues during fetal development. ERβ expression is initiated after ERα during adulthood. ERs regulate cell growth and differentiation in adult tissues [40]. Thus, an appropriate balance between ERα and ERβ is required for normal development of male reproductive tissues. [39]. In male αERKO mice, the testes are smaller than those in wild-type animals and the fertility rates are significantly reduced [41]. Low fertility in αERKO male mice is due to reduced sperm counts and low sperm quality. In contrast, the βERKO males have been shown to produce a sufficient number of sperm to maintain fertility. These findings imply that ERα is more important than ERβ for reproduction system development and sperm maturation in male mice.
Bone development and maintenance in ER knockout mice
Bone plays a role in the support of soft organs and maintains calcium level homeostasis in blood. This tissue is continuously remodeled to maintain a suitable length and density for the body. Calcium, vitamin D, and steroid hormones such as estrogen are critical for bone homeostasis [42,43]. These factors control gene expression in bone cells including osteoblasts, osteoclasts, and osteocytes [44]. In addition, various factors are regulated by estrogen during the formation the bone, which modulate osteoblast differentiation and the development of osteoporosis [45]. Estrogen is known to stimulate the release of growth factors such as insulin-like growth factor 1 (IGF-1) that influence bone resorption. ERKO mice have been very valuable for elucidating the association between mechanisms underlying bone functions and ER signaling [46]. ERα is expressed at low levels in bone. In αERKO mice, bone length and size are significantly decreased compare to normal mice. The bones of male αERKO mice also show a reduction in mineral density [47]. However, skeletons of βERKO mice are phenotypically identical to those of wild-type animals, indicating the low importance of ERα in osteoporosis progression [48].
Cardiovascular tissues and metabolism in ER knockout mice
The rate of cardiovascular system disease for women increases with age after menopause. Although the direct effect of estrogen on the cardiovascular system is unclear, reduced estrogen levels are associated with the disruption of lipid or glucose regulation [49]. The ability of estrogen to decrease cholesterol levels is due to the elimination of low-density lipoprotein (LDL) from blood circulation. In addition, various studies have recently demonstrated that estrogen affects blood vessel structure [5,50]. ERs are expressed in endothelial and smooth muscle cells of vascular tissues, suggesting that ER signaling involves in function of smooth muscle cells. Moreover, ERKO mice possess inherently vascular disease [51]. However, ERα is expressed in tissues involved in glucose and lipid metabolism such as the brain, adipose tissue, liver, and heart [52]. Imbalanced ERα/ERβ ratios may lead to the development of metabolic diseases [53]. αERKO mice are diabetic and insulin resistant. βERKO mice exhibited insulin responsibility and have not excessive fat in the body [54].
Brain and behavior
Estrogen is fundamental for regulating the growth and differentiation of axons and dendrites in the brain [55]. Functions performed by ERα and ERβ in the brain have been explored by studies of αERKO and βERKO mice. Interestingly, previous studies demonstrated differences in localization and expression levels of both ER isoforms. In addition, the results of these studies have shown that the function of ERα seems to be important for reproduction while that of ERβ may affect cognitive processes [5,6,56]. However, the level of ER expression in the brain rapidly changes in response to injury [57]. ERα primarily contributes in masculinization while ERβ contributes to the defeminization of sexual behavior [58]. ERKO females display aggressive behavior and depression. Additionally, estrogen signaling contributes to brain integrity and exerts different effects on behavior [59].
Summary
Estrogen regulates the growth, development, and physiology of the human reproductive system. This hormone also influences neuroendocrine, skeletal, adipose, and cardiovascular systems. Estrogen is an important sex hormone produced primarily by the ovaries in females and testes in males. The biological actions of estrogen are mediated by binding to the ERs in target organs. ERα and ERβ are transcriptional factors that regulate the expression of specific genes in different tissues on a ligand dependent manner. Analysis of ERα and ERβ functions using knockout mouse models has demonstrated the roles of estrogen signaling in different physiological processes. αERKO and βERKO mice are also valuable for examining the effect of ER signaling in specific target organs. Although transgenic mice do not always produce consistent results, these models are useful for evaluating the functions of genes under specific pathological conditions.

 XML Download
XML Download