

 PDF
PDF ePub
ePub Citation
Citation Print
Print


The incidence of hearing loss in humans is substantial, with a frequency of pre-lingual deafness as high as 0.1-0.2% and a similar frequency of post-lingual deafness before the third decade of life [1,2]. In developed nations, ~50% of these cases appear to have a genetic basis. Inherited deafness in humans is genetically heterogeneous, with effects in any one of more than 100 distinct genes likely to be responsible for nonsyndromic hearing loss [1]. Despite the difficulty inherent in the analysis of genetically heterogeneous conditions, there has been dramatic progress in the localization and identification of a large number of genes associated with hearing loss during the past several years [3-5]. Mutations in 15 different genes, including the transmembrane inner ear (TMIE), have been shown to cause nonsyndromic, recessively inherited hearing loss in humans [3]. Recently, loss of function mutations in the Tmie/TMIE gene has been shown to cause deafness in humans (DFNB6). These results indicate that the Tmie/TMIE gene has a conserved, critical role in the auditory system [6,7]. Mouse genetic models provide a valuable approach to identify genes which play a role in inherited human hearing loss, and also offer a useful system to investigate gene function [8]. Identification and analysis of these genes in mice has implicated a diverse array of proteins required in the inner ear during early embryonic development and postnatal maturation of the sensory neuroepithelium, as well as in the adult organ [9]. The Tmie gene has no sequence similarity with other known genes and their functions are presently unclear. Previously our research group reported that the circling mouse is a possible animal model for deafness and it has a 40 kb deletion that includes the Tmie [10,11]. On the basis of these results, circling mice are believed to be an excellent animal model for investigating inner ear abnormalities in humans. However the functional roles of Tmie in the cochlea and other organs such as the liver, lung, kidney and spleen remain unclear. The levels of expression, the distribution and the time course of Tmie have not been defined. Only the cochlear pathology of circling mice and the expression of Tmie protein in various organs of adult rats have been analyzed [10,12]. All these results led us to investigate the expression pattern of Tmie protein in the postnatal developmental stages of C57BL/6J mice.
C57BL/6J mice were used throughout this study. C57BL/6J mice were obtained from the Korea Research Institute of Bioscience and Biotechnology (Deajon, South Korea) and bred in our facilities. The mice were kept in a specific-pathogen free animal care facility and were housed individually in plastic cages (18×30×15 cm) with corn cob bedding. The facility was maintained at 22±2℃ temperatures, in 55±10% relative humidity, and a 12 hr light and 12 hr dark routine cycle was employed. Normal rodent pellet diet (Jeiljedang, Seoul, Korea) and water was supplied ad libitum. The animal study was conducted in accordance with the guidelines and with the approval of the Institutional Animal Care and Use Committee of Hallym University (Hallym-1-51).
Tissue samples (kidney, liver, lung, spleen, and cochlea) were harvested from mice of different age groups (7, 21.42, and 63). We were used western blotting analysis for the expression of Tmie protein in various organs in different age groups. The tissue homogenates (whole cell extract) were prepared by the sonication of these tissues in lysis buffer (50 mM Tris-HCl, pH 6.8, 100 mM DTT, 2% SDS, 10% glycerol). These tissue homogenates (50 µg of the whole cell extract) were boiled for 5 min and then resolved by SDS PAGE electrophoresis (15% gel) runs at 200 V for 60 min in a running buffer (25 mM Tris, 192 mM Glycin, 0.1% SDS). Electrophoretic transfer to a polyvinylidene fluoride transfer membrane (PVDF, Pall Corporation, USA) was then carried out at 100 V for 90 min using the transfer buffer according to the manufacturer's instructions. The membrane was blocked with 5% BSA in PBS at room temperature for 1 h, then incubated for overnight at 4℃ with the anti-Tmie antibody diluted to 1/1,000 in 1% BSA buffer, followed by incubation with horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG (Molecular Probes, USA) for an additional 1 h. The blot was finally developed using Luminol (PerkinElmer Life Sciences, Inc., USA) and then examined with FUJIFILM Luminescent Image Analyzer and Image Reader LAS-1000 Lite software (Fuji Photo Film Co., Ltd, Japan).
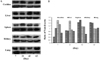
Tmie expression in various mouse organs, including the cochlea, liver, spleen, kidney, and lung, has been identified by using the anti-Tmie antibody which was previously developed by our group [12,13]. A single immunoreactive band for Tmie protein was found in the cochlea, liver, spleen, kidney, and lung tissue samples (Figure 1). The blot was also identified with an anti-β actin antibody to determine the relative level of protein in each lane. The relative expression level of Tmie protein in each organ was determined by the ratio of the intensity of the immunoreactive bands against the anti-Tmie and anti-β actin antibodies. As shown in Figure 1A, the expression level of Tmie protein in the cochlea increased throughout the postnatal developmental stages, indicating that Tmie plays an important role in the development and function of the cochlea. In addition, there was the highest expression level of Tmie protein at 42 days of age in the liver. The expression level of Tmie protein in the spleen and kidney slightly decreased throughout the postnatal developmental stages. In the lung, Tmie protein expression increased throughout the study period. Figure 1B show ratios of the intensities of the immunoreactive bands against anti-Tmie and anti-β actin antibodies. The order of Tmie expression level was cochlea, kidney, liver, lung, and spleen at 7 days after birth (cochlea 1.6, spleen 0.4). This pattern of Tmie expression level changed with later stages of mice. At 63 days after birth, the order of Tmie expression level was spleen, liver, lung, cochlea, and kidney. The Tmie expression level in several organs of adult rats has been reported previously [15]. Order of Tmie expression level in adult rats was liver, cochlea, lung, spleen, and kidney.
To know features of Tmie protein, we analyzed Tmie protein using UniProt analysis (Figure 2). Tmie protein has a signal peptide (1-28 AA), extracellular region (29-58 AA), transmembrane region (59-79 AA), cytoplasmic region (80-153 AA) and lycine-rich domain (124-153 AA). Tmie protein is need for correct development of stereocilia bundles in the cochlea [16]. The mature Tmie protein is localized in the plasma membrane [13]. From features of Tmie protein by UniProt analysis and previous results, Tmie protein may reside within an internal membrane compartment and function in vesicle trafficking.
There have been a few reports that deafness genes have been involved with cancer and liver transplantation patients having hearing impairment side effects [17,18]. As a primary step to reveal the deafness proteins related with other organs, we examined the level of Tmie protein expression in these organs. Taken together, these results show that Tmie protein could be important and that it could play a role in the function of these organs on the basis of the needs of each particular organ. More functional studies including localization of Tmie protein in liver, spleen, lung, and kidney should are needed to understand the function of Tmie in these organs.

 XML Download
XML Download