

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Atherosclerosis, which has been linked to abnormalities in nitric oxide (NO) signaling, is a chronic systemic disease of the vasculature with an inflammatory component. NO is produced by nitric oxide synthase (NOS), which has three isoforms: neuronal NOS (nNOS), endothelial NOS (eNOS), and inducible NOS (iNOS) [1]. Both eNOS and nNOS are constitutively expressed. In contrast, iNOS expression is inducible by activated macrophages as an immune reaction. During infection, iNOS produces large quantities of NO [2]. iNOS knockout mice decreased diet-induced atherosclerosis [3]. apolipoprotein E (ApoE)/iNOS double knockout mice showed significantly smaller atherosclerotic lesions compared to ApoE knockout mice, suggesting that reduction in iNOS-mediated oxidative stress may protect lesion formation in double knockout mice [4].
Numerous studies have demonstrated that activation of the angiotensin II type 1 (AT1) receptor plays an important role in the pathogenesis of inflammatory diseases. AT1 receptor blockers reduced lipopolysaccharide (LPS)-induced upregulation of iNOS gene and protein expression and iNOS activity, which decreased the general peripheral inflammatory reaction to LPS [5]. LPS-stimulated glomerular iNOS expression was enhanced in diabetic nephropathy, and the activation of angiotensin II may play a role in this enhancement [6]. Olmesartan, a new AT1 receptor blocker, significantly decreased the expression of iNOS in myocardial tissue with a decrease in myocardial inflammation [7]. Losartan, an AT1 receptor blocker, reduced iNOS expression and reestablished the redox status, thus reducing early markers of diabetic retinopathy in a model of diabetes and hypertension [8].
Flavonoids are polyphenolic compounds that are widely distributed in plants, and their consumption might be associated with decreased risks for some chronic degenerative diseases in humans [9]. Epidemiological research in Japan has shown that green tea consumption protects against coronary atherosclerosis [10]. Recent evidence has shown that interleukin-1β (IL-1β)-induced expression of iNOS was markedly inhibited in human chondrocytes pretreated with epigallocatechin-3-gallate (EGCG), a flavanol that is enriched in green tea [11]. EGCG reduced IL-1β and interferon-γ (IFN-γ)-induced NO production, and reduced levels of iNOS mRNA and iNOS protein in RINm5F cells [12]. Ex vivo analysis of β-islets showed that EGCG down-regulates expression of iNOS [13] induced by multiple low doses of streptozotocin. EGCG inhibited effects induced by ultraviolet B (UVB) including activation and translocation of NF-κB, expression of iNOS mRNA and generation of NO, indicating that EGCG protects against UVB-induced skin damage [14]. However, the effect of EGCG on the expression of iNOS, a major risk factor for vascular inflammation, remains unknown. Accordingly, we investigated this knowledge gap using human umbilical vein endothelial cells (HUVECs). This could be clinically important because specific inhibitors of iNOS expression (such as EGCG) might be helpful for the treatment of cardiovascular diseases such as atherosclerosis. We hypothesized that EGCG reduces the expression of iNOS and reactive oxygen species (ROS) that is induced by angiotensin II in HUVECs.
Materials and Methods
Materials
All antibodies for Western blotting were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). The culture medium was obtained from Invitrogen (Carlsbad, CA, USA). Angiotensin II, EGCG and other reagents were purchased from Sigma-Aldrich (St. Louis, MO, USA), unless otherwise specified.
Cell culture
HUVECs were obtained from Clonetics (Walkersville, MD, USA). They were grown in medium 199 containing 0.1 mg/mL heparin, 25 µg/mL endothelial cell growth factor (Biomedical Technologies, Stoughton, MA, USA), 2 mM L-glutamine, 100 U/mL penicillin G, 100 µg/mL streptomycin and 20% fetal bovine serum (FBS). The medium was renewed every two days until confluence, when cells were subcultured at a 1:3 ratio and then cultured in an atmosphere of 95% air and 5% CO2 at 37℃.
Western blot analysis
HUVEC cultures were starved for 12 h and treated with the desired drugs for the desired times. Cells were lysed in ice-cold buffer (20 mM Tris-HCl pH 7.4, 1% Triton X-100, 150 mM NaCl, 1 mM EDTA, 1 mM EGTA, 2.5 mM sodium pyrophosphate, 1 mM β-glycerol phosphate, 1 mM Na3VO4, 1 mM PMSF and 1 µg/mL leupeptin). The lysates were sonicated and centrifuged (12,000 rpm, 20 min). The protein concentration was measured by the Bradford method. Equal amounts of protein (10 µg) were run on 12% SDS-PAGE and blotted onto polyvinylidene difluoride membranes. These were incubated with rabbit polyclonal antibodies (1:100) against iNOS. Secondary anti-rabbit antibodies and enhanced chemiluminescence (ECL) Plus reagent kits (Amersham, Little Chalfont, Buckinghamshire, UK) were used for detection. Membranes were subsequently exposed to ECL hyperfilms.
Detection of ROS
Cells were starved in phenol red-free M199 medium containing 1% FBS for 12 h and stimulated with angiotensin II and 2',7'-dichlorofluorescein diacetate for 2 h. Fluorescence signals were quantified (Molecular Devices, Sunnyvale, CA, USA).
Statistical analysis
Results are shown as the means±SEM from at least three independent experiments. Statistical significance between the means was assessed by one-way ANOVA followed by Tukey's multiple comparison test; P<0.05 was taken as statistically significant.
Go to : 
Results
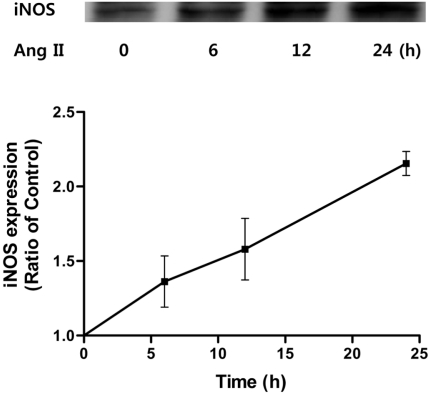
Angiotensin II increased the levels of iNOS in HUVECs
To determine whether expression of iNOS, a risk factor for vascular inflammation, is affected by angiotensin II, HUVECs were treated with angiotensin II. Angiotensin II (100 nM) increased the expression of iNOS in a time-dependent manner (Figure 1) causing iNOS levels to increase for the next 24 h. Thus, angiotensin II increases protein levels of vascular endothelial iNOS.
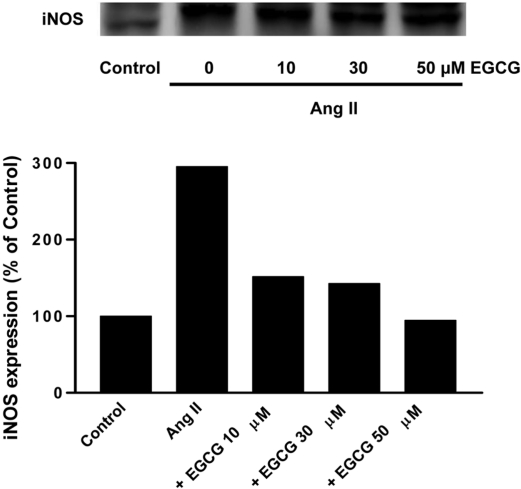
Effect of EGCG on the expression of iNOS induced by angiotensin II in HUVECs
To determine whether angiotensin II-stimulated iNOS expression is affected by EGCG, HUVECs were pretreated for 0.5 h with 10, 30, 50 µM EGCG prior to treatment with angiotensin II (100 nM) for 24 h. Increasing concentrations of EGCG inhibited Ang II-induced iNOS expression (Figure 2) Thus, EGCG, a major catechin obtained from green tea leaf, decreases the protein level of iNOS in a concentration-dependent manner.
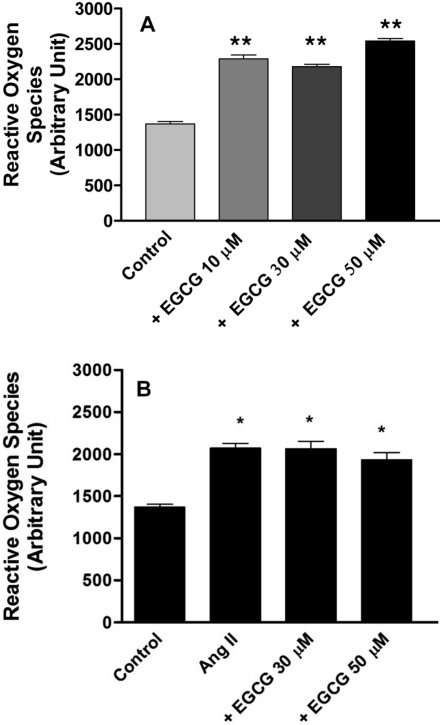
Effect of EGCG on the production of ROS in HUVECs
To determine whether ROS production is affected by EGCG, HUVECs were pretreated with EGCG. EGCG (10, 30 and 50 µM) increased ROS production compared with control (no treatment), (Figure 3A). Angiotensin II (100 nM) increased ROS production compared with control and this was not prevented by 30 or 50 µM EGCG (Figure 3B). These data suggest that EGCG decreases the protein level of iNOS without affecting ROS.
Go to :
Discussion
Inflammation is an important event in the development of vascular diseases such as hypertension, atherosclerosis, and restenosis [15]. Recent knockout studies on iNOS have clarified the role of NO in inflammatory responses [16]. Plasma levels of the proinflammatory cytokines tumour necrosis factor-α (TNF-α), IL-6, and IL-1β were also significantly reduced in iNOS-knockout mice in comparison with control wild-type mice [17]. Expression of iNOS in the endothelium may be a more important mediator of vascular dysfunction than expression of iNOS in the adventitia [18]. Excess NO resulting from iNOS activation was important in modulating vascular permeability during sepsis [19]. iNOS was partially responsible for pulmonary oxidative and nitrosative stress reactions in a murine model [20]. A selective iNOS inhibitor, ONO1714, retarded the development of atherosclerosis [21] induced by a high-cholesterol diet. Increased iNOS activity was found in circulating neutrophils of septic shock patients compared to healthy volunteers [22].
Angiotensin II, a peptide hormone, also plays an important role in atherosclerosis. Ang II, through activation of NF-κB-mediated pro-inflammatory genes, promoted vascular inflammation, leading to acceleration of atherosclerosis [23]. Inhibition of NF-κB by the antioxidant pyrrolidine dithiocarbamate (PDTC) markedly reduced inflammation and iNOS expression in a double transgenic rat model, in which rats transgenic for the human angiotensinogen and renin genes are crossed [24]. Angiotensin II via AT1 receptor increased the accumulation of neutrophils and iNOS expression and played a significant role in mediating inflammation in gastric mucosa exposed to ischemia-reperfusion [25]. Irbesartan, an AT1 receptor antagonist, attenuated atherosclerosis. This effect was partly related to the inhibition of oxidative stress and inflammatory signal transduction pathways in high cholesterol-diet ApoE knockout mice [26]. Increased formation of angiotensin II and iNOS in infiltrated macrophages and medial smooth muscle cells (SMCs) might play important roles in the development and progression of human coronary atherosclerosis [27].
Catechin, a flavanol derived from green tea, possesses antioxidant, antiangiogenesis and antiproliferation activities during the prevention and treatment of cardiovascular diseases [28]. EGCG decreased the risk of cardiovascular disease by reducing inflammatory markers in rats fed an atherogenic diet [29]. In the EGCG groups, iNOS expression was reduced in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-treated mice [30]. EGCG activated an adenosine 5'-monophosphate-activated protein kinase and blocked LPS/IFN-γ-induced iNOS expression, supernatant NO and IL-6 in mesangial cells from MRL/lpr lupus-like mice [31]. Pretreatment of NHBE cells with EGCG resulted in a significant down-regulation of NF-κB-regulated proteins such as cyclin D1, matrix metalloproteinase-9 (MMP-9), IL-8 and iNOS [32]. In the ventral prostate, EGCG significantly reduced cell proliferation, induced apoptosis, and decreased androgen receptors, insulin-like growth factor-1 (IGF-1), IGF-1 receptor, phospho-extracellular signal-regulated kinases 1 and 2, cyclooxygenase-2 (COX-2), and iNOS [33]. Administration of EGCG significantly suppressed iNOS transgene induction by IFN-γ and LPS [34].
In this study, we investigated whether EGCG reduces angiotensin II-mediated iNOS expression in HUVECs. Angiotensin II increased the levels of iNOS (Figure 1). EGCG prevented the iNOS expression induced by angiotensin II in a concentration-dependent manner (Figure 2). This novel finding for the effects of green tea/EGCG is complemented by findings that resveratrol, an important antioxidant found in grapes and wine, diminished IFN-γ-induced levels of iNOS protein, attenuated iNOS mRNA levels, and inhibited IFN-γ-induced promoter activity of the iNOS gene [35]. Resveratrol inhibited NO production by NO scavenging and down-regulation of iNOS expression in RAW 264.7 cells [36]. Moreover, quercetin, a dietary flavonoid, decreased mRNA expression of inducible enzymes such as COX-2 and iNOS, and thus has protective effects against the oxidized low-density lipoproteins (ox-LDL) induced inflammation in peripheral blood mononuclear cells [37]. Quercetin inhibited myocardial ischemia-reperfusion-induced iNOS mRNA and protein expression in rabbit [38]. Sulforaphane, mainly derived from cruciferous vegetables, inhibited the expression of iNOS that had been stimulated by LPS in HUVECs [39]. Together, these results show that flavonoids have a common inhibitory effect on iNOS expression in a number of different cell types in a variety of conditions.
To understand the mechanisms for EGCG attenuation of iNOS expression by angiotensin II, we examined that ROS attenuation in HUVECs induced by EGCG led to an inhibition of iNOS expression. Our data demonstrate that EGCG increases ROS and does not affect ROS increased by angiotensin II (Figure 3). EGCG triggered ROS resulted in chondrosarcoma cell death [40]. However, EGCG decreased the angiotensin II-induced increase of O2- and ROS in vascular smooth muscle cells [41]. Therefore, interference with ROS production by EGCG may be different in different cell types and under a variety of conditions. Alternatively, overproduction of ROS by EGCG might inhibit the expression of iNOS in HUVECs.
In conclusion, the anti-atherosclerotic activity of EGCG is likely not associated with decreased ROS production, although there may be some benefit through inhibiting the expression of iNOS induced by angiotensin II.
Go to :
 XML Download
XML Download