

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Uterine adenocarcinomas have been reported in most domestic species but are considered rare except in rabbits and cows [1]. The prevalence of uterine adenocarcinoma in rabbits after 5 years of age has been reported to be 79% [2]. Because rabbits are induced ovulators that are often housed individually, laboratory rabbit and possibly many pet rabbits are under almost constant estrogen stimulation. Virgin rats have a similar high incidence of endometrial adenocarcinomas [3]. Cats, another species that exhibit induced ovulation, may also be subjected to long periods of unopposed estrogen stimulation, but the frequent practice of neutering queens, and the tendency to periodically breed or induce ovulation in sexually intact queens, may explain the low prevalence of endometrial adenocarcinoma in cats [4]. In women, postmenopausal hormone replacement therapy, estrogen in particular, is considered a risk factor for endometrial adenocarcinoma; both estrogen-dependent and higher-grade estrogen-independent adenocarcinomas are recognized [5].
Viruses cause cancer in amphibians, fish, fowl, rodents, cats, cattle, and subhuman primates, and if humans were to be exempt from such a general biological phenomenon, it would be a circumstance unparalleled in the history of parasitism. In fact, evidence is accumulating that some human cancers are virally induced. For example, Burkitt's lymphoma, nasopharyngeal carcinoma, cervical cancer, and Kaposi's sarcoma are thought to be caused by herpesviruses, which are able to remain latent in the host for many years [6]. Feline leukemia virus (FeLV) is an exogenous retrovirus, belonging to the genus Gammaretrovirus, which infects domestic cats and, sporadically, wild cats as well [7,8]. FeLV causes a wide range of proliferative diseases in cats, including lymphoid and myeloid leukemia [9].
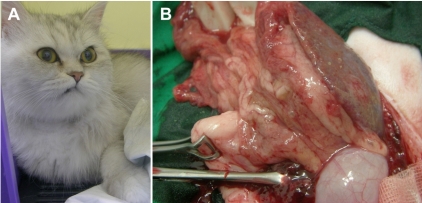
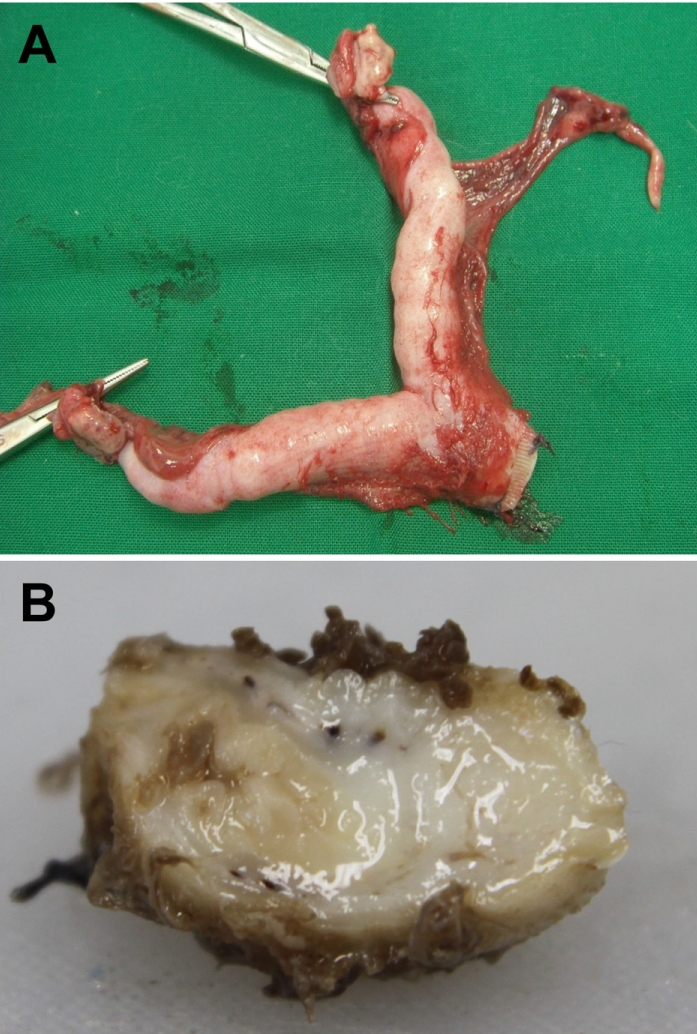
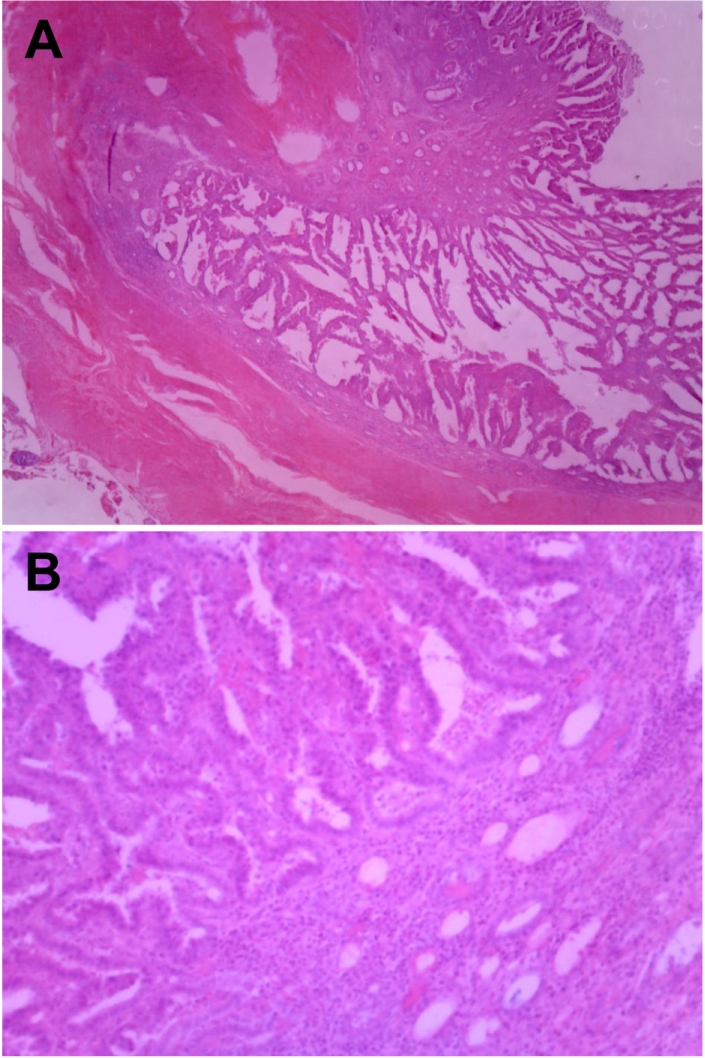
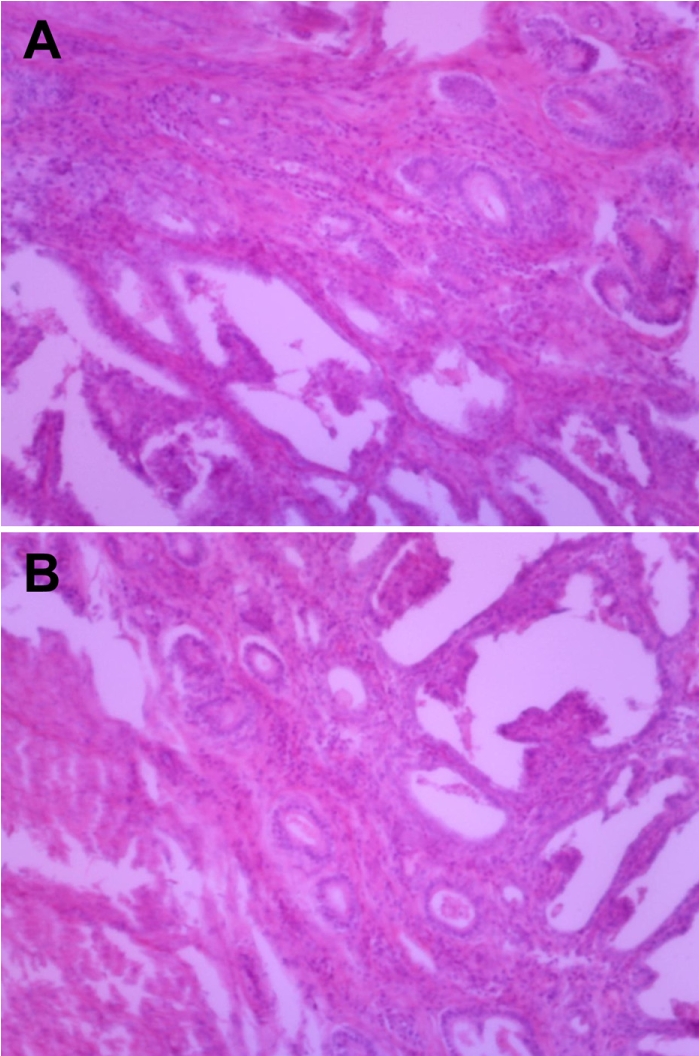
Feline endometrial adenocarcinomas are uncommon malignant neoplasms that have to date been poorly characterized. In this study, a case of uterine adenocarcinoma case is described in a Persian cat with FeLV infection. At the time of presentation, the cat was a 2-year-old female Persian chinchilla (Figure 1A). She presented with vaginal discharge and interrupted vomiting, and was given a health examination. Abdominal sonography and radiography demonstrated abnormal enlargement of the uterus in the abdominal cavity. The blood analysis revealed neutrophilia and lymphopenia. The concentrations of progesterone and estrogen in blood were evaluated and the results revealed normal ranges. The cat underwent surgical ovariohystectomy. During the laparotomy, thickening and inflammatory lesions of the omentum and spleen were observed (Figure 1B). The results of bacterial culture with omentum and spleen samples were negative for pathogenic bacteria. The ovariouterus and the omentum were removed surgically, submitted to gross examination, and trimmed. Cross-sections of the uterine wall revealed a thickening of uterine horn (Figure 2). Additionally, yellowish contents were observed in the lumen of the uterus. The trimmed tissues were fixed in 10% neutral buffered formalin and embedded in paraffin. Four-micrometer sections were made and stained with hematoxylin and eosin (H&E) for histopathological examination. The histopathological analysis revealed adenocarcinoma in the uterus and omentum (Figure 3), but no pathological changes were observed in the ovary. The adenocarcinoma had metastasized to the omentum, resulting in thickening and the formation of inflammatory lesions (Figure 4). Based on these histopathological findings, the case was diagnosed as uterine adenocarcinoma with abdominal metastasis. The cat made a complete recovery following an ovariohysterectomy and chemotherapy.
The blood samples were tested for FeLV, feline infectious peritonitis (FIP), feline immunodeficiency virus (FIV), feline panleukopenia virus (FPV), feline herpesvirus (FHV), feline calicivirus (FCV), heart worm, feline chlamydia, Toxoplasma, Babesia spp., Ehrlichia spp., Haemobartonella felis, Rickettsia spp., and Brucella spp. using a polymerase chain reaction (PCR) assay. The following primer sequences were used for PCR amplification: 5'-CTACCCCAAAATTTAGCCAGCTACT-3' and 5'-AAGACCCCCGAACTAGGTCTTC-3' for the amplification of the 468-bp segment of FeLV [10]; 5'-TAATGCCATACACGAA CCAGCT-3' and 5'-GTGCTAGATTTGTCTTCGGACACC-3' for the amplification of the 295-bp segment of FIP [11]; 5'- CCACAATATGTAGCACTTGACC-3' and 5'-GGGTACTTTCTG GCTTAAGGTG-3' for the amplification of the 583-bp segment of FIV [12]; 5'-CATTGGGCTTACCACCATTT-3' and 5'-GGTGC ACTATAACCAACCTCAGC-3' for the amplification of the 172- bp segment of FPV [13]; 5'-CGGGAAAATCCAGTACGAGT-3' and 5'-AGGAAGAGTTCGGCGGTATT-3' for the amplification of the 383-bp segment of FHV [14]; 5'-TTCGGCCTTTTGTG TTCC-3' and 5'-TTGAGAATTGAACACATCAATAGATC-3' for the amplification of the 673-bp segment of FCV [15]; 5'- AGTGCGAATTGCAGACGCATTGAG-3' and 5'-AGCGGGTA ATCACGACTGAGTTGA-3' for the amplification of the 542- bp segment of heartworm [16]; 5'-ATGAAAAAACTCTTGA AATCGG-3' and 5'-CAAGATTTTCTAGACTTCATTTTGTT-3' for the amplification of the 1069-bp segment of feline chlamydia [15]; 5'-AGTTTAGGAAGCAATCTGAAAGCACAT C-3' and 5'-GATTTGCATTCAAGAAGCGTGATAGTAT-3' for the amplification of the 529-bp segment of Toxoplasma [16]; 5'-ATAACCGTGCTAATTGTAGG-3' and 5'-TGTTATTTCTTGT CACTACC-3' for the amplification of the 327-bp segment of Babesia spp. [17]; 5'-GGAATTCAGAGTTGGATCATGGCTC AG-3' and 5'-CGGGATCCCGAGTTTGCCGGGACTTCTTCT- 3' for the amplification of the 492-bp segment of Ehrlichia spp. [18]; 5-AGCAGCAGTAGGGAATCTTCCAC-3' and '5- TGCACCACCTGTCACCTCGATAAC-3 for the amplification of the 674-bp segment of Haemobartonella felis [19]; 5'- AGAGTTTGATCCTGGCTCAGAAC-3' and 5'-CCTACGGCTA CCTTGTTACGACTT-3' for the amplification of the 356-bp segment of Rickettsia spp. [20]; 5'-TCGAGCGCCCGCA AGGGG-3' and 5'-AACCATAGTGTCTCCACTAA-3' for the amplification of the 905-bp segment of Brucella spp. [21].
Genomic DNA was isolated from blood using an AccuPrep Genomic DNA extraction kit (Bioneer, Daejeon, Korea) according to the manufacturer's instructions. The DNA was eluted in Tris-EDTA buffer (pH8.0), and an aliquot was used for PCR amplification. All DNA samples were stored at -20℃ until the PCR assays were performed. The template DNA (50 ng) and 20 pmol of each primer were added to a PCR mixture tube (AccuPower PCR PreMix; Bioneer) containing 2.5 U of Taq DNA polymerase, 250 µM each deoxynucleoside triphosphate, 10 mM Tris-HCl (pH8.3), 40 mM KCl, 1.5 mM MgCl2, and the gel loading dye. The final volume was adjusted to 20 µL with distilled water. The reaction mixture was subjected to denaturation at 95℃ for 2 min, followed by 40 cycles of 95℃ for 30 sec (denaturation), 54℃ for 30 sec (annealing), and 72℃ for 30 sec (extension), and a final extension step of 72℃ for 5 min, and samples were kept at 4℃ until analysis. Reactions were conducted using My Genie 32 Thermal Block PCR (Bioneer). Eight microliters of each sample were mixed with 2 µL of loading buffer, and electrophoretically separated on 1.2% agarose gels stained with 0.5 µg/mL ethidium bromide. DNA bands were observed under ultraviolet light.
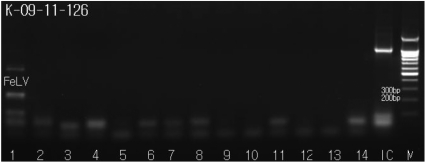
The PCR-amplified samples were evaluated for the presence of PCR amplicons corresponding to FeLV, FIP, FIV, FPV, FHV, FCV, Heart worm, Feline Chlamydia, Toxoplasma, Babesia spp., Ehrlichia spp., Haemobartonella felis, Rickettsia spp. and Brucella spp. We were able to visualize bands that were 529 base pairs in length, corresponding to the FeLV amplicon (Figure 5); other feline pathogens tested negative in the PCR assay (Figure 5).
Significant changes in the structure of the endometrium are associated with degeneration of luminal epithelium, cystic endometrial hyperplasia, pyometra, and adenocarcinoma [22]. Endometrial tumors of the uterus are rare in domestic animals [23]. Adenocarcinoma of the uterus is very rare in animals, in marked contrast to the prevalence of this tumor in human females [23]. The basic mechanisms associated with these changes are poorly understood. Prolonged exposure to megestrol acetate has been associated with endometrial hyperplasia and pyometra in domestic cats, and progestin contraceptives may have similar effects on zoo felids [24,25]. Megestrol acetate is a derivative of progesterone and used commonly both in contraception and hormone replacement therapy [26]. Based on available clinical and epidemiological data, it has been long suspected that tumors of the canine female genital tract develop under the influence of ovarian hormones [22]. In this case, however, the animal had not been given a progesterone derivative such as megestrol acetate.
FeLV is an exogenous retrovirus causing several diseases in domestic cats, including tumors of most hematopoietic cells, aplastic anemia, myeloproliferative disorders, and immunosuppression [27]. The FeLVs that primarily cause T-cell tumors in cats are transmitted in a horizontal manner [28], particularly through saliva [29]. The T-cell tropism of these agents is observed even when the virus infects human cells [30]. Although many free-roaming cats become persistently infected with FeLV, only a small fraction develops leukemia or lymphoma. A large number develop lethal opportunistic infections, particularly coronavirus-induced peritonitis [31]. FeLV causes lymphopenia, depressed cell-mediated immune responses, thymic atrophy, and depressed humoral responses to T-cell dependent antigens despite the presence of hypergammaglobulinemia [31].
In this study, the cat had uterine adenocarcinoma without pathognomic ovarian lesions or significant hormonal imbalances. The cat also suffered from FeLV infection and lymphopenia. We believe that the uterine adenocarcinoma in this case was probably related to immunogenic depression by FeLV infection, although we cannot rule out the contributions of other factors, such as old age, to the induction of this tumor. To the best of our knowledge, this is the first report of feline uterine adenocarcinoma with feline leukemia virus infection.
 XML Download
XML Download