

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Deafness is the most common form of sensory impairment in humans. Over the last decade, enormous progress has been made in the discovery of genes involved in hearing and deafness, as well as in the identification of many new loci and specific mutations that cause heritable deafness and vestibular disorders [1]. There has been a particularly tremendous escalation in the localization and identification of genes for nonsyndromic hearing impairment. Inherited deafness in humans is genetically heterogeneous, with effects in any one of more than 100 distinct genes likely to be responsible for nonsyndromic hearing loss [2]. The incidence of hearing loss in humans is high, with a frequency of prelingual deafness as high as 0.1-0.2% and a similar frequency of post-lingual deafness before the third decade of life. Researchers now believe that more than 60% of congenital deafness cases in developed nations are caused by genetic factors [3]. Despite difficulties in analysis of genetically heterogeneous conditions, there has been dramatic progress in the localization and identification of a large number of genes associated with hearing loss during the past several years. The causes of nonsyndromic deafness are complex. Researchers have identified more than 30 genes that, when mutated, may cause nonsyndromic deafness; however, some of these genes have not been fully characterized.
Recently, loss-of-function mutations in the transmembrane inner ear (Tmie) gene have been shown to cause deafness in mice and humans [4-9]. These results indicate that the Tmie gene has a critical role in the auditory system. Previously our research group has reported that circling mice are a possible animal model for deafness. These mice have a 40-kb genomic deletion, including the Tmie gene [6,10-11].
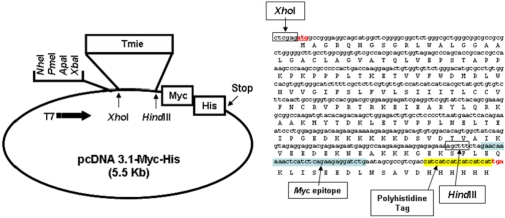
In order to understand the subcellular localization and functional relationships of the Tmie protein, we developed a stable cell line for expressing Tmie protein. The cells (HEK293) were transfected with the Tmie expressing vector [pcDNA 3.1-Myc-His (5.5 Kb)-Tmie] and the expression of Myc-tagged Tmie was confirmed by Western blot analysis and immunostaining analysis using anti-Myc and anti-Tmie antibodies. Our results provide an excellent model for studying the synthesis and localization of Tmie protein.
Materials and Methods
Construction of a Tmie expression vector
A Tmie expressing vector [pcDNA 3.1-Myc-His (5.5 Kb)-Tmie] was used [12]. Briefly, mouse Tmie cDNA was amplified by polymerase chain reaction (PCR) using the primer sets: 5'-CTGGACTCTCAGGACCTGCA-3' and 5'-TCAGGAAGCC GCCCTCATTT-3'. The amplified PCR product was ligated into the XhoI and HindIII sites of mammalian expression system vector pcDNA3.1-Myc-His (Invitrogen, Grand Island, NY, USA) to yield the Tmie expression construct pcDNA 3.1-Tmie-Myc-His. DNA sequencing was used to verify the nucleotide sequences of the Tmie expression vector.
Construction of a stable cell line
The human embryonic kidney cell line (HEK293) was obtained from the American type culture collection. HEK293 was maintained with Dulbecco's modified Eagles medium containing 10% fetal bovine serum, 25 mM HEPES, 100 U/mL, penicillin and 100 µg/mL streptomycin. Cells were cultured at 37℃ at 95% of air and 5% CO2 atmosphere. To generate a stable cell line, we transfected 5 µg of the Tmie expressing vector [pcDNA 3.1-Myc-His (5.5 Kb)-Tmie] which confers neomycin resistance into HEK293 cells, using FuGENE 6 transfection reagent (Roche, Mannheim, Germany) according to manufacturer instruction, Two days after transfection, cells were selected in 500 µg/mL G418 (Duchefa Bioch, Haarlem, Netherlands) for 2 weeks.
Western blot analysis
Cells (5×106) were lysed in lysis buffer (50 mM Tris-HCl pH7.4, 150 mM NaCl, 1 mM EDTA, 0.1% SDS, 1% Triton X-100, 1 mM PMSF). Clarified lysates were resolved on 12% SDS-polyacrylamide gels and then transferred to polyvinylidene fluoride transfer membranes (Millipore, Billerica, MA, USA). The membranes were blocked in Tris-buffered saline containing 0.05% Tween 20 and 2% bovine serum albumin for 1 hr at room temperature. Membranes were incubated with an anti-Myc antibody (1:1,000, Cell Signaling Technology, Danvers, MA, USA) for 2 h. Immunoreactive proteins were detected using horseradish peroxidase-conjugated secondary antibody (1:500, Jackson ImmunoResearch Laboratories, West Grove, PA, USA) and enhanced Chemiluminescence reagent (Amersham Pharmacia Biotech, Piscataway, NJ, USA).
Immunofluorescence staining
For immunofluorescence staining, the stable cells were cultured on glass coverslips in 12-well plates for 24 h in order to detect Tmie localization. Cells were fixed with 4% paraformaldehyde, permeabilized with 0.1% Triton X-100, incubated with antibodies against Myc epitope (1:200), and then incubated with Alexa Flour 633 conjugated goat anti-mouse IgG (Molecular Probes, Grand Island, NY, USA). Nuclei were stained with DAPI. Coverslips were mounted in fluoromount-G (Southern Biotechnology Associates, Birmingham, AL, USA). The mounted samples were scanned with an LSM 510 (Carl Zeiss, Jena, Germany). Several positive clones were expanded and analyzed in this study.
Results
Stable expression of Myc-tagged Tmie in HEK 293 cells
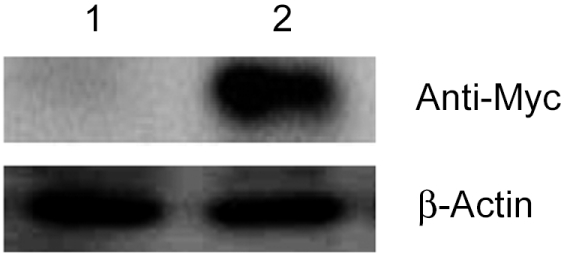
Because we aimed to generate cell lines that stably express Tmie, we transfected the vector containing Tmie and a neomycin resistance gene (Figure 1). In order to monitor expression of Tmie, we introduced a Myc epitope into the end of Tmie gene. The vector sequences were confirmed by an automatic sequencer. Thereafter HEK 293 cells were selected with G418 for stable expression of the transgenes. The Tmie protein expression level of the established cell lines was determined by western blotting analysis as described in methods. The Tmie expression level in the established cell lines was monitored through several passages by Western blot analysis. Stable transgene expression was observed with different levels (Figure 2). No band was detected in untransfected (empty vector) cells.
Subcellular localization of Myc-tagged Tmie
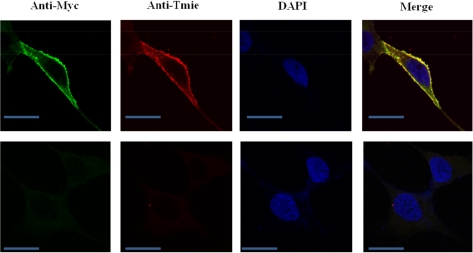
Along with analyzing the expression levels, we also studied the subcellular localization of Tmie in the established cell lines. The subcellular localization of Myc-tagged Tmie was studied by immunofluorescent staining followed by confocal microscopy. Detailed subcellular localization analysis performed with HEK293 cells clearly demonstrated that the Myc-tagged Tmie is expressed predominantly in the plasma membrane (Figure 3).
Discussion
Inherited hearing loss is genetically heterogeneous and is caused by mutations in gene encoding proteins responsible for a variety of processes, including the maintenance of hair cell structure, neuronal innervations, and trafficking. Recently, a high-throughput detection method of the mutations responsible for hearing loss was established [13]. It has been found that Tmie is responsible for nonsyndromic hearing loss in humans [7-9]. The Tmie protein has a signal peptide and at least one transmembrane domain [5]. According to the proposed structure of Tmie, the mature protein may be localized in the plasma membrane and serve as a site of interaction for other molecules through its highly charged C-terminal domain. Several papers suggest that the Tmie protein may require maturation of sensory cells as well as the normal development of the maintenance of stereocilia [5,14-16].
To examine the subcellular localization of the Tmie protein we generated a cell line that expresses the Tmie protein. The Tmie protein expression in HEK293-Tmie cells was monitored by using immunostaining analysis and the localization was observed through confocal microscopy. From this study we concluded that the Tmie protein is mostly present in the cellular membrane, which suggests that Tmie protein may play an important role in signal trafficking between the cells or in signal transduction in the auditory system. Tmie protein probably joins with other proteins while transducting signals in the auditory system.
We predict that the Tmie protein may play an important functional role in cell metabolism and signal transduction. However, more specific study is needed to understand its functional role in adjacent cells. It remains to be determined what specific role Tmie plays in hair cell development and specifically, what advantages its localization and membrane association provide. However the data provided in this study offer a solid foundation from which these questions can better be addressed.
In summary, we established a Tmie-expressing cell line on HEK 293 cells, and the expression of Tmie was confirmed by both western blotting and confocal microscopy. Considering the shortcomings involved in functional analysis of Tmie, our method ensures stable and high expression of the protein, thus providing an excellent model for further research into its biological effects. Further examination of the impact of Tmie and its protein will provide insight into the pathways that are critical for normal maturation of sensory cell function. Analysis of Tmie expression and localization will also help to define the molecular mechanism of hearing loss in humans with defects at the DFNB6 locus, and will provide a system to investigate potential therapies that rescue sensory cells and preserve auditory function.
 XML Download
XML Download