

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Peroxiredoxins (Prxs) are a 24-kDa peroxidase that belongs to an antioxidant enzyme family. The cysteine (Cys) residue on this protein is the primary site of oxidation and acts as an electron donor for the reduction of peroxides [1,2]. Six isoforms of mammalian Prxs (I-VI) were identified with similar immunological properties and amino acid sequences [3,4]. The distribution of the Prxs isoforms in the human brain was found to vary. Prx I was primarily expressed in astrocytes, while Prx II was expressed in neurons of various region including the cerebral cortex, hippocampus, cerebellum, basal ganglia, substantia and spinal cord. Moreover, they were differentially located within cells. Prx I, II and VI were mainly distributed to the cytosol, but Prx III and V were largely present in the organelles and Prx IV was secreted into the extracellular region [5,6].
Of the six isoforms, Prx I was predominantly expressed in various type of tumors and functioned as an anti-apoptotic protein for tumor cells proliferation and survival [2]. In addition, several studies have found that Prx I was tightly correlated with neurodegenerative disease [7,8]. The expression level of Prx I was not significantly altered in Down syndrome (DS), Alzheimer's disease (AD) and Pick's disease (PD) when compared to the control [7]. Especially, AD which was showed the massive accumulation of extracellular Aβ-42 peptides produced by γ-secretase composing of four subunits and the hyperphosphorylation of Tau proteins has been received great interest from scientists [9,10]. The Aβ-resistance PC12 cell line showed higher expression levels of multiple Prxs isoforms than that of the control cells with reduced cysteine oxidation. Furthermore, an increase in Aβ-resistant was induced by transfection of wild type Prx I in PC12 cells and rat primary hippocampal neurons [8]. However, the effects of Prx I on the expression of the γ-secretase complex on AD have not yet been studied and are unknown. Therefore, in this study, we investigated whether Prx I could regulate the expression of the γ-secretase complex, which causes AD, in cells overexpressing Prx I and an AD animal model.
Materials and Methods
Care and use of animals
The animal model for AD was produced by the microinjection of the human Pen-2 gene, a key regulator of γ-secretase complex, into the pronucleus of fertilized eggs as described previously [11]. All animal experimental procedures were approved by the Institutional Animal Care and Use Committee (IACUC) at the Pusan National University (Approval No.: PNU-2010-000220). All mice were supplied by the breeding center of Korea FDA facility and were housed in cages under a strict light cycle (lights on at 06:00 h and off at 18:00 h) and constant temperature of 23±1℃. In addition, all mice were provided a standard irradiated chow diet (Purina Mills, St. Louis, MO, USA) ad libitum and maintained in a specific pathogen free (SPF) state.
Reverse transcription-polymerase chain reaction (RT-PCR) analysis in tissue of Tg mice
For the preparation of total RNA, tissues frozen in liquid nitrogen were chopped with scissors and homogenized in a RNA-Bee™ solution (Tel-Test, Austin, TX, USA). The isolated RNA was then quantified using an Ultraspec 1000 system (Amersham Pharmacia Biotech, Buckinghamshire, UK). To characterize the expression of transgenes, RT-PCR was conducted using 5 µg of total RNA from each of the tissue samples. 500 ng of Oligo-dT primer (Invitrogen, Carlsbad, CA, USA) was annealed for 10 min at 70℃. Complementary DNA, which was utilized as a template for further amplification, was synthesized via the addition of dATP, dCTP, dGTP and dTTP, as well as 200 units of reverse transcriptase. In these reactions, 10 pmoles of the sense and antisense primers were added, and the reaction mixtures were subjected to 30 cycles of amplification. Amplification was conducted in the aforementioned thermal cycler under the following conditions: 30 sec at 94℃, 30 sec at 62℃ and 45 sec at 72℃. In each case, minus-RT controls were included to distinguish between the DNA and RNA products. This experiment was repeated three times, and the relative differences in RNA quantity were also reproducibly observed in the three experiments. The sequences of the sense and antisense primers for Pen-2 were 5'-GCTAT GAACC TGGAG CGAGT G-3' and 5'-GAAGG AGAGG TAGTC CCCAA GG-3', Prx I were 5'-GCGCT AGCGG ACTGC TGATA GGAAG ATGTC-3' and 5'-GCCTC GAGCA GCGCT CACTT CTGCT TGGAG-3', Prx VI were 5'-GCGCT AGCCT TGTTC TCAGC GTCAC CACTG-3' and 5'-GCCTC GAGCC AGTAC TGGAT GTGCA GATGC AG-3', β-actin were 5'-TGGAA TCCTG TGGCA TCCAT GAAAC-3' and 5'-TAAAA CGCAG CTCAG TAACA GTCCG-3', respectively. Finally, the levels of each Pen-2, Prx I and Prx VI RT-PCR product were quantified using the aforementioned electrophoresis documentation and analysis system on a 1% agarose gel.
Western blotting
SH-SY5Y cells harvested from 100 mm-diameter culture dishes and the tissue from non-Tg and NSE/hPen-2 Tg mice were solubilized and homogenized with 1% nonidet P-40 in 150 mM NaCl, 10 mM Tris HCl (pH 7.5), and 1 mM EDTA, and supplemented with a protein inhibitor mixture (Roche, Basel, Switzerland). From 15 to 30 µg of protein was separated by electrophoresis on a 10% polyacrylamide gel for 2 h and the resolved species were transferred to a nitrocellulose membrane by electroblotting for 2 h. The membrane was incubated with primary anti-human Pen-2 antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA, 1:1,000 dilution), anti-PS-2 antibody (Cell Signaling Technology, Boston, MA, USA, 1:1,000 dilution), anti-APP antibody (Sigma-Aldrich, St. Louis, MO, USA, 1:4,000 dilution), anti-APH-1 antibody (Sigma-Aldrich, 1:1,000 dilution), anti-NCT antibody (Cell Signaling Technology, 1:1000 dilution), anti-Prx I antibody (Abcam, Cambridge, UK, 1:1000 dilution), anti-Prx VI antibody (Abcam, 1:1000 dilution) or anti-actin antibody (Sigma-Aldrich, 1:3,000 dilution) overnight at 4℃. Each membrane was washed with buffer (137 mM NaCl, 2.7 mM KCl, 10 mM NaHPO4, and 0.05% Tween-20) and incubated with a 1:1,000 dilution of horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG at room temperature for 2 h. The membrane blots were developed using Enhanced Chemiluminescence Reagent Plus kit (Amersham).
Perfusion and immunohistochemcal analysis
Brain perfusion and immunohistochemical analyses was performed as previously described [12,13]. Briefly, mice were anaesthetised with Zoletil 50 (Virbac, Carros cedex, France) and transcardially perfused with 1X PBS followed by 4% formaldehyde to effectively remove the blood and fix the brain tissue. After perfusion, each mouse brain was isolated from the skull and fixed overnight in formaldehyde. Each brain was dehydrated and embedded in paraffin. A series of brain sections (10 µm) were cut from paraffin-embedded tissue using a Leica microtome (Leica Microsystems, Bannockbrun, IL, USA). For immunohistochemical analysis, these sections were de-paraffinized with xylene, rehydrated and pretreated for 30 min at room temperature with PBS blocking buffer containing 10% goat serum. The sections were then incubated with the anti-Aβ-42 antibody (Invitrogen), at a dilution of 1:100 in PBS blocking buffer. The antigen-antibody complexes were visualized with biotinylated secondary antibody (goat anti-rabbit)-conjugated HRP streptavidin (Histostain-Plus Kit; Zymed, South San Francisco, CA, USA), at a dilution of 1:1,500 in PBS blocking buffer. The Aβ-42 peptides were detected using stable 3,3'-diaminobenzidine (DAB; Invitrogen) and observed using a model BX50F-3 optical microscope (Olympus, Tokyo, Japan).
Gene constructions
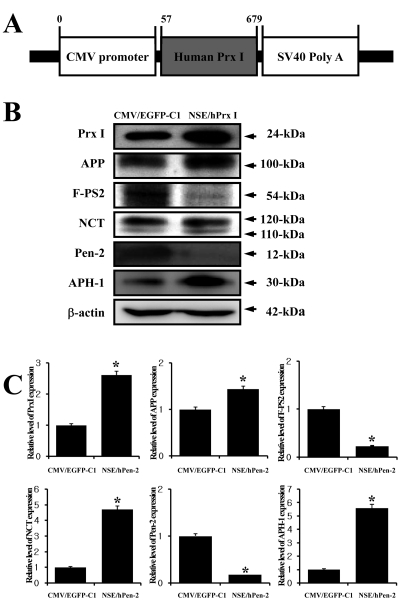
The CMV/hPrx I plasmid, which harbors hPrx I under the control of the CMV promoter, was constructed. Briefly, the hPrx I gene (GenBank accession No. BC021683.1) was amplified by the PCR using a full-length RNA isolated from NCI-H460 human carcinomal cells. The primers used for the amplification were hPrx I sense primer, 5'-GCgctagc GGACT GCTGA TAGGA AGATG TC-3' (italic letter, NheI site; ATG, start codon; capital letter, hPrx I sense primer corresponding to nucleotides 57-78 of hPrx I) and the hPrx I anti-sense primer, 5'-GCctcgag CAGCG CTCAC TTCTG CTTGG AG-3' (italic letter, XhoI site; the capital letter corresponds to nucleotides 658-679 of hPrx I). The primers used contained a recognition sequence for the NheI and XhoI enzymes at the 5' and 3' end of the PCR products, respectively. The amplified hPrx I was 622-bp in length, and the product was cloned into the T cloning vector (pGEM-T; Promega, Madison, WI, USA). Sequence analysis was conducted in order to confirm whether or not the cloned hPrx I sequence was identical to hPrx I cDNA, and the resulting sequence was aligned with the NCBI sequence database, using the BLAST program to identify their corresponding hPrx I gene. The hPrx I cDNA fragment isolated using NheI and XhoI digestion was inserted into the pEGFP-C1 vector in order to construct the CMV/hPrx I recombinant vector. Finally, the CMV/hPrx I plasmid was subjected to enzyme digestion and automatic sequence analysis to confirm their sequence and orientation (Figure 3A).
Cell cultures and DNA transfection
NCI-H460 cells and SH-SY5Y cells were purchased from the Korean Cell Line Bank (Seoul, Korea). Cells were grown in monolayers in RPMI 1640 (Hyclone Co., Logan, UT, USA) supplemented with 10% fetal bovine serum (FBS; Invitrogen) and antibiotics (100 unit/mL penicillin and 100 µg/mL streptomycin; Invitrogen) during incubation at 37℃ in a humidified incubator containing 5% CO2 in air. All other chemicals were purchased from Sigma-Aldrich. To overexpress the Prx I protein in neuroblastoma cells, the cells were transfected with the CMV/hPrx I plasmid using the Lipofectamine™ Reagent (Invitrogen). SH-SY5Y cells were seeded at a density of 2×106 cells per 100 mm-diameter dish. After reaching 80-90% confluence, a plasmid-lipofectamine mixture containing 20 µg of DNA (per dish) was added, and the cells were incubated for an additional 5 h at 37℃ in OptiMEM medium. The cells were then cultured in fresh serum-free medium for 24 h and were used for Western blot and flow cytometry analyses.
Results
Accumulation of Aβ-42 peptides in the brain of NSE/hPen-2 Tg mice
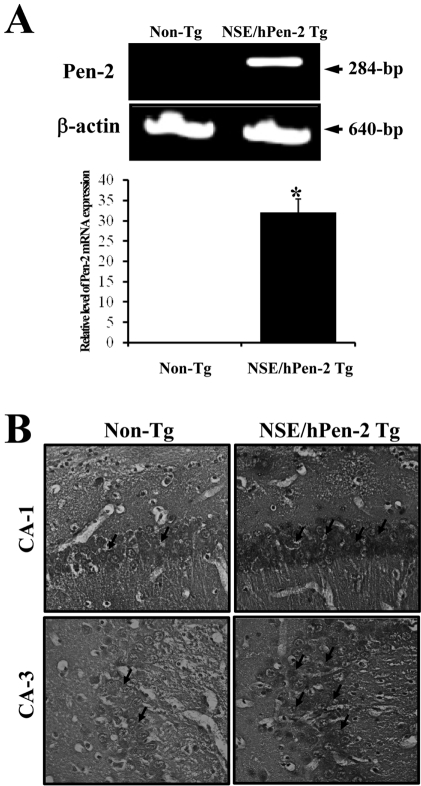
Remarkable accumulation of Aβ-42 peptides is one of most important features in the animal model for AD [12]. Pen-2 expression was first detected using RT-PCR analysis. The mRNA expression of Pen-2 was dramatically higher in the brain of NSE/hPen-2 Tg mice (Figure 1A). In order to investigate the accumulation of Aβ-42 peptides in the brain of NSE/hPen-2 Tg mice, the level of Aβ-42 peptides was detected by immunohistochemical analysis. NSE/hPen-2 Tg mice contained higher levels of Aβ-42 peptides in the hippocampus of brain than the Non-Tg mice (Figure 1B). Therefore, the NSE/hPen-2 Tg mice used in this study had the distinct features of AD.
Change of Prx I and VI expression in the brain of NSE/hPen-2 Tg mice
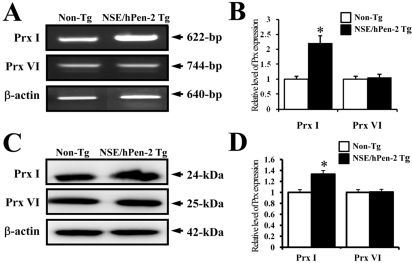
Next, to investigate whether accumulation of Aβ-42 peptides could alter Prx I and VI expression, the Prx I and VI mRNA and protein levels were detected in the brain of Non-Tg mice and NSE/hPen-2 Tg mice. The expression level of Prx I mRNA was significantly higher in NSE/hPen-2 Tg mice compared to Non-Tg mice, while the expression level of Prx VI was not different between the two groups (Figure 2A). Also, the protein and mRNA levels of both enzymes were very similar, although the ratio was different in each analysis (Figure 2A). These results suggested that accumulation of Aβ-42 peptides induced by Pen-2 overexpression may upregulate Prx I but not Prx VI.
Effect of Prx I overexpression on the expression of γ-secretase complex
To study the effect of Prx I on the expression of the γ-secretase complex, SH-SY5Y neuroblastoma cells were transfected with the CMV/hPrx I plasmid and the expression level of each component from γ-secretase was detected using the specific antibody. The expression level of Prx I was successfully increased in the CMV/hPrx I transfectants when compared to the vector transfectants. In addition, the expression level of APP, which is a substrate of γ-secretase, was slightly higher in the CMV/hPrx I transfectants than the vector transfectants. Of the different components of the γ-secretase complex, the expression level of key factors including PS-2 and Pen-2 was lower in the CMV/hPrx I transfectants when compared to the vector transfectants, while the expression level of the assistant factor (APH-1 and NCT) was higher in the CMV/hPrx I transfectants. These results suggested that Prx I may down-regulate PS-2 and Pen-2 and up-regulate APH-1 and NCT of the γ-secretase complex.
Discussion
Prx I is a member of the 2-Cys subfamily and is considered a candidate therapy target of lung adenocarcinoma since overexpression of Prx I has been observed in various cancer cell lines [14,15]. However, most of studies have focused on the role of Prx I in the development, progression and drug resistance of cancer [14,16]. Only a few studies have examined the correlation between Prx I and neurodegenerative diseases. In this study, we demonstrated that Prx I could regulate the expression of key molecules responsible for neurodegenerative diseases.
Especially, the expression level of PS-1 protein, a component of the γ-secretase complex, was strongly correlated with oxidative stress. In both the cells lysate and lipid raft fraction of neuroblastoma SH-SY5Y cells, a significant increase in PS-1 protein expression was observed in cells subjected to oxidative stress; however, under the same conditions, no changes in the expression of NCT, APH-1, Pen-2 or BACE-1 were observed. Furthermore, this increase in PS-1 protein expression was prevented by co-treatment with an antioxidant [17]. In this study, an antioxidant condition was induced by transfection of CMV/hPrx I vector, which altered the expression level of γ-secretase complex components. As shown in Fig. 3, the expression level of the NCT and APH-1 protein was significantly increase by the overexpression of Prx I. However, PS-2 and Pen-2, which is a coregulator of γ-secretase activity, was higher under the same condition. Therefore, our results showed that the expression level of the γ-secretase complex components may be altered by oxidative stress conditions as well as the overexpression of antioxidant enzymes.
Several studies had provided conflicting results on the expression levels of Prxs in the brain of AD. In a study that examined Prxs expression in three neurodegenerative diseases, Down syndrome (DS), AD and Pick's disease (PD), the expression level of Prx I under the disease condition were reported to not be significantly different from the control. However, the expression level of Prx II was found to be dramatically higher in the frontal cortex for these three diseases, where Prx III expression was lower in same region of DS and PD [7]. Similar results were observed in Tg mice overexpression Aβ-42 peptides and amyloid binding alcohol dehydrogenase. Proteomic analysis of the brain from these mice showed that the Prx II level was higher in the AD mice model [18]. However, another study reported that the expression level of Prx I was higher in post-morten human AD cortex tissue than in age-matched control [8]. In our study, the expression level of Prx I was significantly higher in Tg mice overexpressing Pen-2 gene compare to non-Tg mice, while the level of Prx VI was the same between these two groups. Therefore, our results suggested that Prx I may be correlated with the pathological condition of AD.
In this study, we investigated the correlation between Prx I and the pathogenesis of AD. These results showed that the accumulation of Aβ pepetides induced the expression of Prx I, but not Prx VI. Also, the overexpression of Prx I by transfection induced an increase in NCT and APH-1 expression and a decrease in PS-2 and Pen-2 expression.
 XML Download
XML Download