

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Chemicals known to have a potential to disrupt the endocrine or hormone system in humans and wild animals by affecting endocrine signals are classified as endocrine disrupting chemicals (EDCs) [1]. Originally, EDCs are defined by the U.S. Environmental Protection Agency (EPA) as exogenous agents that interfere with synthesis, secretion, transport, metabolism, binding, action or elimination of natural blood-borne hormones that are present in the body and are responsible for homeostasis, reproduction and developmental process [2]. EDCs are contained in a variety of chemical classes, including drugs, pesticides, compounds used in the plastic industry, consumer products, industrial by-products and pollutants, and even some naturally produced botanical chemicals. They have been remarkably increasing in the environment with the rapid growth in these industries [3] and therefore, human and animals are sustainedly exposed to a wide variety of EDCs [4]. Health effects attributed to EDCs include a range of reproductive problems such as reduced fertility, male and female reproductive tract abnormalities, skewed male/female sex ratios, loss of fetus, and menstrual problems [5]. Specifically, EDCs have been generally supposed to affect sex steroid hormone-controlled organs and tissues in the reproductive tract through nuclear hormone receptors, such as estrogen receptors (ERs), androgen receptors (ARs), progesterone receptors (PRs) and thyroid receptors (ThRs) [2,6]. Among these receptors, estrogen-mimicking activities through ERs of some EDCs were first proved to cause endocrine disruption in the 1950s and these EDCs are generally called as 'xenoestrogens'. Xenoestrogens mean "foreign" estrogens having estrogenic effects on a living organism even though they differ chemically from the estrogenic substances internally produced by the endocrine system of the organism. Usually, xenoestrogens may act as false messengers and mainly disrupt the reproduction process related with endogeneous estrogen. Diethylstilbestrol (DES), alkylphenols (APs), bisphenol A (BPA), polychlorinated biphenyls (PCBs) and phthalates are representative xenoestrogens having similar structures to 17β-estradiol (E2) in EDCs. Starting in the 1990s, extra mechanisms related with other hormonal system were found in the endocrine disruption of industrial compounds. For instance, the disruption of thyroid hormone transport and the altered androgen antagonism were identified in the actions of other EDCs [7]. As a result of the endocrine disruption via several pathways, EDCs may cause hormone imbalance, early puberty, brain and behavior problems, impaired immune functions and various cancers [8]. When it comes to cancer, E2 itself is also known as one of risk factors in tumorigenesis of the estrogen-dependent organs, such as endometrium, breast, and ovary [4,6]. Therefore, xenoestrogens as estrogen mimics may increase the potential for cancer lesions by transforming normal patterns of tissue and interrupting dominant regulatory mechanisms in the estrogen dependent organs. Meanwhile, unlike most chemically synthesized EDCs, some phytoestrogens, which are produced as secondary metabolites in some plants and act as estrogen in animals [9,10], are known to have cancer chemopreventive effects. According to previous reports, high level diets of phytoestrogen may have protective effects on estrogen-related diseases, such as prostate and breast cancers [9,11]. In this respect, this review will focus on not only the xenoestrogenic properties of various EDCs and their carcinogenic potentials but the preventive effects of phytoestrogens on estrogen-dependant cancers substantiated from the in vivo studies. Until now, the roles of EDCs in carcinogenesis have been reviwed mainly focusing on the causation of specific cancers like mammary cancer by well known EDCs such as DES and BPA [7,12,13]. But, this review will be anticipated to provide a comprehensive point of view to understand cancer causation and prevention by various EDCs.
EDCs have estrogen-like properties via ERs signaling pathway
17β-Estradiol
E2 is an endogenous female sex steroid hormone and very important in the estrous cycle and the development of reproductive organs in human and animals. As a hormone binding to its receptor triggers a number of events, E2 preferentially binds to ERs. ERs are expressed in the estrogen-dependent organs such as endometrium, breast and ovary [14,15]. There are two forms of ERs, ERα and ERβ, which play unique physiological roles depending on the tissue and cell types [13]. ERs are activated by binding E2 and in turn modulate the expression of many related genes via ERs signaling pathways [16]. The genomic pathway is a well known classical pathway of ERs signaling and mediates target gene regulation via direct binding to an estrogen response element (ERE), which is a specific DNA sequence within the promoter, of an estrogen-ER complex [17-19]. The ERE/receptor complex then recruits other transcription factors that are responsible for the transcription of downstream target genes and finally induces protein synthesis. As a result, this pathway promotes the development of female secondary sexual characteristics and is involved in the thickening of the endometrium and in regulating the menstrual cycle.
Endocrine disrupting chemicals
Generally, as EDCs have similar structures to E2, they may exhibit an estrogenic activity through the interaction with ERs like E2. Though the binding affinities of EDCs are relatively low compared to E2, they can activate ERs and regulate cellular functions induced by ERs [3]. For instance, by way of the genomic pathway, EDCs mediate the expression of many genes through direct interaction with ERE or through interaction with transcription factors, including members of activator protein-1 (AP-1), nuclear factor-κB (NF-κB), signal transducer and activator of transcription (STATs), and families of specificity protein (SP-1), especially aryl hydrocarbon receptor (AhR), or via modification of estrogen metabolism [2,20]. In the previouse studies, the xenoestrogenic effects and the risks of endocrine disruption of diverse chemicals have been identified. The disruption effects of some well-known EDCs on estrogen signaling pathways by their xenoestrogenic properties are as in the following.
First, DES is a potent synthetic estrogen and a drug once prescribed during pregnancy to prevent miscarriages or premature deliveries from the 1940s to 1970s [21,22] and now classified as a representative harmful xenoestrogen by interacting with ERs with very high affinity and influencing genomic signaling [22,23]. Alkylphenols (APs) are common chemicals used to make alkylphenol ethoxylate surfactants in household detergents and personal care products and widely used as plasticizers and chemical stabilizers in various industries [24]. Nonylphenol (NP) and octylphenol (OP) are typical forms of APs and easily discharged through sewage disposal works [25]. APs bind ERα and prompt ligand-dependent gene expression via ERE promoter. Also, APs can reduce steroidgenesis and disrupt endocrine system via ER indirect pathways [26,27]. While they have been used in industry for over 40 years, the European Union has implemented sales and use restrictions on certain applications in which nonylphenols are used because of their alleged "toxicity, persistence, and the liability to bioaccumulate" [28]. BPA is one of the EDCs that has been the most completely studied. Because BPA is used in many consumer goods such as baby bottles, beverage cans, dental sealants and other many kinds of plastics, it has the highest opportunity to be exposed to the environment [29]. A 2011 study that investigated the number of chemicals pregnant women are exposed to in the U.S. found BPA in 96% of women [30]. The binding affinity of BPA to ERs is lower than E2 or DES, but BPA has been interacting with the ERs in a way that is completely unique in all known classes of ER ligands. BPA functions as a xenoestrogen by binding strongly to estrogen-related receptor γ (ERR-γ) which is an orphan receptor (endogenous ligand unknown) that behaves as a constitutive activator of transcription [31]. Also, BPA has been exhibited to interact with the ERs as a partial agonist with both AF-1 (ligand-independent activation) and AF-2 (ligand-dependent activation) [32,33].
Dichlorodiphenyl-trichloroethane (DDT) is one of the most well-known synthetic pesticides and is most commonly known among the EDCs. DDT is mainly metabolized to 1,1-dichloro-2,2-bis(4-chlorophenyl) ethylene (DDE) and their effects have been widely studied [34,35]. DDE can activate multiple receptors such as ERα, AR, and PR and transcription factors and intrude on estrogen biosynthesis by increasing an aromatase activity, the key enzyme for catalyzing the rate-limiting step in the transformation of androgens into estrogen [36].
PCBs were widely used as dielectric and coolant fluids in transformers, capacitors, and electric motors. As omnipresent and persistant in the environment, they may cause the health problems in humans and wildlife such as disorders in reproductive, nervous, immune and endocrine system [37]. Concerns about the toxicity of PCBs are largely based on the similarity in chemical structure and toxic mode of action with dioxin. As one of EDCs, many parts of their effects are known to be ER-mediated by acting as estrogen mimics or as estrogen antagonists [37,38].
Phthalates are a class of chemicals widely known as plasticizers in personal-care products, children's toys, pharmaceuticals, food products, and textiles and so on. Dibutylphthalte (DBP), butyl benzyl phthalate (BBP), and di(2-ethylhexyl)phthalate (DEHP) are typical forms of phthalates. Recent studies report that several phthalates affect the developmental process in animals such as a significant increase in the number of aberrations in chromosome separations in oocytes at anaphase in various aquatic organisms and the alterations in gonadal development and spermatogenesis in amphibians [39-42] and also cause the developmental problems in human like a shortened anogenital distance among baby boys [43]. In addition, the exposure of phthalates was associated with mental, motor, and behavioral development problem in children and was harmful for formation of genital tract in both male and female [44,45]. In ligand binding assays, phthalates were shown to weakly compete with E2 for binding ER and to have weak ER-mediated activity [46]. In addition, fetal male rats exposed to DBP or DEHP ostensibly showed the deformities in androgen-dependent tissues in sexual differentiation in vivo by a non-receptor mediated mechanism [47].
Go to : 
EDCs have potential effects of carcinogenesis
Estrogen-dependent cancers
Regarding the carcinogenicity, previous studies of EDCs have described that developmental exposure to xenoestrogens can transform normal patterns of tissue, and these mutations may interrupt dominant regulatory mechanisms and improve the potential for cancer lesions [48,49]. Especially, the xenoestogens can be risk factors to estrogen dependant cancers such as breast, ovary and prostate cancers. Breast cancer is a multifactorial disease of humans and other mammals and is highly dependent on sex and the level of sex hormones such as estrogen and progesterone [50]. Therefore, breast cancer has relevance to exposure of exogeneous EDCs as well as endogeneous hormone level. Many epidemiological studies searched the relation between the exposure to EDCs and breast cancer [51,52]. For instance, women exposed to DES during pregnancy showed a modest increase in the incidence of breast cancer risks. Daughters born after in utero exposure to DES have shown the more development of breast cancer after the age of 40 when compared to unexposed women of the same age [53,54]. Ovarian cancer is a cancer arising from the ovary of women and most ovarian cancers are classified as "epithelial" believed to arise from the surface of the ovary. For ERs in normal condition, ERβ is highly expressed in granulose cells of the ovary, whereas ERα is expressed at relative low levels in thecal and interstitial cells. Several studies have reported that ERα is overexpressed in ovarian cancer compared to normal tissue and promotes the proliferation of cancer cells [6,55]. Prostate cancer is also a hormone-responsive cancer developing in the male reproductive system. Although prostate cancer is highly dependent on adrogens, estrogens also play a role in prostate cancer pathogenesis. ERα and ERβ are originally expressed in different cell types in prostates; ERα is mainly expressed in stroma and ERβ in epithelium. These two receptors are thought to play different and opposing roles in prostate cancer with ERα having proliferative properties and ERβ having anti-proliferative properties [56]. But other studies suggest that ERβ is highly expressed in the prostate and may be a susceptible target for treating prostate cancer lesions [57,58].
Carcinogenic effects of EDCs in in vivo animal models
Generally, prenatal exposure to EDCs can change the development of the mouse mammary gland and increase the susceptibility to further carcinogenic exposure in the adults. For example, mammary specific alterations due to early life BPA exposure have been identified by several studies, in which administrating low concentrations (25-1,000 µg/kg/day) of BPA via subcutaneously implanted osmotic pumps have shown the alterations to mammary gland morphology in adult animals following perinatal exposure to BPA in mice [59]. In rats, perinatal exposure to environmentally relevant doses of BPA enhanced the number of intraductal hyperplasias that occurs in adulthood, altered mammary gland morphogenesis at high doses, and induced precancerous lesions and the development of carcinomas in situ [60]. Exposure to BPA for 50 days in feeding period showed an increased number of dimetylbenz [a]anthracene (DMBA)-induced tumors per rat and a decreased latency period compared to animals not exposed to BPA [61]. At puberty, an enhanced sensitivity to estrogen was shown in the mammary glands of animals exposed to BPA, which led to increased duct lateral branching [62]. Additionally, ERα and PR overexpression was shown in the endometrial epithelium and lamina propria of adult mice that were exposed to BPA in uterus [63]. BPA was also shown to enhance the expression of vascular endothelial growth factor (VEGF) in the immature rat at 37 mg/kg of the lowest active dose [64]. When VEGF is overexpressed, it can contribute to cancer to grow and metastasize. Also, the vascular permeability in uterus of ovariectomized mice was increased by BPA [65].
For dioxins, its metabolite, 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD), is reported to be strongly involved in carcinogenesis. Prenatal exposure to TCDD had shown a significant interference in mammary gland differentiation and morphogenesis as a greater number of terminal end buds [66]. In mice, TCDD blocks estrogen-induced responses in several tissues [67]. Exposure of rats to the carcinogen, dimetylbenz[a]anthracene (DMBA), at puberty enhanced the tumor occurrence and reduced the latency period as compared to animals not exposed to TCDD [68,69].
DEHP, a typical phthalate, is known as a peroxisome proliferator and a anticipated human carcinogen because it induces 5- to 10-fold dose-related increase in liver tumor proliferation in both sexes of rats and mice [70,71]. DEHP was also identified to have estrogenic response in uterotrophic and vaginal cornification assay [72]. Also, in immature ovariectomized SD rats, DEHP caused the significant growth in uterine weight by oral injection (20, 200 or 2,000 mg/kg) [73].
Chemopreventive effects of phytoestrogens in in vivo animal models
Phytoestrogens are poly-phenolic non-steroidal substances such as isoflavonodis, flavonoids, stilbenes and lignans. Many phytoestrogens are shown to have estrogenic effects by selectivity binding ERβ in competitive ER binding and transcriptional assay [74]. Several animal studies showed that phytoestrogens contended with estrogen in ERs binding and regulated normal ERs action in target tissues [75,76]. As the most examined phytoestrogen, genistein is an isoflavone present in much higher quantities in human diet and in serum when compared to endogenous estrogen, E2. Many studies have shown that genistein is able to binding to both of the ERs, has 20-fold expanded affinity to ERβ as compared with ERα and has more mighty transcriptional activity in cells transfected with ERβ [77-79]. In the respect of cancer prevention, genistein has been shown to inhibit cell growth in breast and prostate cancers by regulating genes responsible for the cell proliferation, cell cycle, apoptosis, and transcriptional regulations [12]. In the molecular level, exposure of genistein reduced mRNA level of transforming growth factor-α (TGF-α) and epidermal growth factor (EGF), and also reduced the protein level of epidermal growth factor receptor (EGFR) and vascular endothelial growth factor receptor 2 (VEGF-R2) in the mammary gland [80,81]. As another well-known phytoestrogen, resveratrol (3,5,4'-trihydroxy-trans-stilbene), is a naturally appearing phytoalexin present mainly in grapes and therefore occurrs high levels in red wine [82,83]. This is a compound generated by plants in reaction to environmental stress or pathogen attack and also demonstrated to have chemopreventive capabilites [75]. This is structurally similar to DES and can bind to both ERα and ERβ. Also, resveratrol reduced E2 stimulated cell growth and constrained transcription of PR in breast cancer cell line [12,84]. But, the antagonist activity of resveratrol is only for ERα and not for ERβ. In mammary carcinogenesis, subcutaneous injections of reveratrol (0, 10 or 100 mg/kg) significantly decreased NMU-induced tumor incidence and diversity [83,85]. Also, other studies showed that in the developing tumors of rats treated with resveratrol, the expressions of COX-2 and matrix metalloproteinase-9 (MMP-9) proteins had significantly decreased [86,87]. Additionally, it was reported that in a mammary tumorigenesis, resveratrol (1, 3 or 5 mg/kg) reduced tumor size and angiogenesis by increasing the apoptotic quotient in mice and rats [88,89]. While many studies support the protective properties of phytoestrogens on cancer formation, some studies insist their tumor progression activities. For instance, it was reported that dietary genistein stimulated tumor growth and antagonized the cytotoxic effects of Tamoxifen and the aromatase inhibitor, Letrozole [90]. Resveratrol also showed detrimental effects on mammary tumor progression [88] or no effect on the same cancer in another study [91]. Although there is a controversy for these mixed results, epidemiological evidences that individuals consuming a diet high in phytoestrogens have a reduced risk of developing various cancers and substantial experimental outcoms are still convincing of the chemopreventive effects of phytoestrogens.
Go to :
Conclusions
Nowadays, the chemicals classified as EDCs are rapidly increasing in environment and therefore, human and animals have been continually exposed to the risks of EDCs. Among diverse EDCs, as xenoestrogens are structurally similar to E2, an endogeneous estrogen, and act as false messengers, they may disrupt the reproduction process related with endogeneous estrogen. Chemically synthetic EDCs such as BPA, DDT, PCB and phthalate are shown to act as estrogen mimics or as antagonists by affecting ERs, ERα and ERβ, and further disturb various cellular functions induced by ERs as shown in Figure 1. As a result, these EDCs may cause hormone imbalance, early puberty, reproduction problems, impaired immune functions and various cancers. In the respect of cancer, many studies of EDCs using in vivo mouse models have described that the exposure to xenoestrogens can transform normal patterns of tissue, interrupt dominant regulatory mechanisms and increase the carcinogenic potential for estrogen-dependant cancers such as breast, ovary, endometrium and prostate cancers. On the other hand, some EDCs have protective effects against estrogen-related diseases. They are mostly phytoestrogens such as genistein and resveratrol and were shown to inhibit cell growth in breast, ovary and prostate cancers by inducing cell cycle arrest, apoptosis, and metastasis suppression via ER-related pathways or non ER-related pathways. Nevertheless, as other studies suggest that phytoestrogens also have tumorigenic properties like other EDCs, the chemopreventive effects of phytoestrogens on estrogen-dependant cancers are still debating.
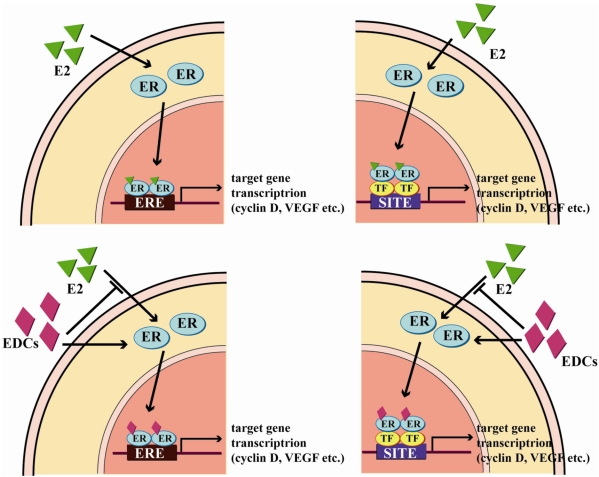 | Figure 1Genomic pathways of estrogen and EDCs in ER-dependent signaling mechanisms. Estrogen and EDCs can lead to cell proliferation in the genomic pathway in which ER dimers directly bind to EREs following ligands binding or ERs interact with other transcription factors (TFs), including AP-1, by action of transcription factor cross-talk. EDCs can compete with estrogen in ERs binding and induce estrogenic effects.
|
The studies of EDCs investigated in this article sufficiently substantiate that EDCs can be sufficient risk factors in various reproduction problems and diseases including specific cancers, but this assertion is somewhat controversal. This is mainly about the validity of dosage of EDCs used in laboratory experiments [92,93]. This is mainly about the validity of dosage of EDCs used in laboratory experiments. Critics argue that the concentrations of chemicals in the experiments are relatively high and that their actual amounts in the environment are too low to cause an endocrine disrupting effect. But, others insist that high dosage effects are appropriate because of EDCs' accumulating properties in living bodies and the environment and high frequency of exposure to EDCs [94]. Therefore, it is prerequisite to validate the effective dosage of chemicals before elucidating their endocrine disruption effects and risk potentials in many disorders. In addition, the health effects of EDCs identified in in vivo studies using animal models should be verified in huamn and other animals. Despite the debate and need for validation, the potential risks of EDCs in endocrine disruption and carcinogenesis are considerbly noteworthy issues in industrialized societies.
Go to :
 XML Download
XML Download