

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Oral and pharyngeal cancer is the sixth most common cancer world-wide and two-thirds of these cases occur in developing countries (Parkin et al, 2005). Despite advances in cancer diagnosis and treatment, there has been little improvement in the 5-year survival rate of oral cancer patients in the last few decades. Because of this, various approaches have been used in the clinical treatment of oral cancer patients in the last three decades, from non-targeted chemotherapy to highly targeted pharmacological inhibitors and specific monoclonal antibodies (Hamakawa et al, 2008; Scully and Bagan, 2009). Although targeted therapies yield better outcomes than non-targeted therapies, frequent treatment failure suggests the need for new strategies for this disease, including new drugs or targets.
Rheum undulatum L. (RL), a well-known traditional Chinese medicine, has been widely used for thousands of years in China for the treatment of many diseases (He et al, 2009). In particular, the root of the species is widely used as a purgative and anti-inflammatory agent in East Asia (Chang et al, 1996; Kuo et al, 2001; Yu et al, 2008). It has been also reported that RL is used traditionally in Korea for the treatment of dental disease (Kim et al, 2010). Recently, it was found that RL contains many anthraquinones and their glycosides, including aloe-emodin, rhein, emodin, and chrysophanol, which are thought to be the major active components for inhibiting tumor growth in various cancer cells (Cha et al, 2005; Huang et al, 2006; Huang et al, 2007; Dorsey and Kao, 2007). However, there has been no study performed to investigate the effect of RL on the growth of human oral cancer cells.
In the present study, we examined the effect of the hexane extract of Rheum undulatum L. (HERL) on the growth of oral cancer cells and the molecular targets for HERL-induced apoptosis. The results demonstrated that HERL induced apoptotic cell death to inhibit the proliferation of oral cancer cells and the down-regulation of Sp1 and survivin mediated the anti-cancer activity of HERL.
Materials and Methods
Reagents
The antibodies for poly(ADP-ribose) polymerase (PARP) and survivin were obtained from BD Biosciences (San Jose, CA, USA) and Cell Signaling Technology (Danvers, MA, USA), respectively. Antibodies for Sp1 and actin were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). 4'-6-Diamidino-2-phenylindole (DAPI) and propidium iodide (PI) were acquired from Sigma-Aldrich (St. Louis, MO, USA). HERL was kindly supplied by professor Jeon JG (Jeonbuk National University, Jeonju, Korea).
Cell culture and chemical treatment
The HN22 and SCC15 human oral cancer cell lines were obtained from the School of Dentistry, Dankook University (Cheonan, Korea). The cells were cultured in Dulbecco's modified essential medium (DMEM) containing 10% fetal bovine serum (FBS) and 100 units/mL penicillin and 100 µg/mL streptomycin (WelGENE, Daegu, Korea) in a humid atmosphere of 5% CO2. Equal numbers of cells were seeded and allowed to attach overnight. The cells were treated with 0.1% NaCl or HERL (20, 40 or 60 µg/mL) diluted in DMEM with 5% FBS for 48 h.
MTS assay
The effects of HERL on cell viability were estimated using the CellTiter 96 Aqueous One Solution Cell Proliferation Assay Kit (Promega, Madison, WI, USA) according to the manufacturer's instructions. The cells were seeded in 96-well plates and incubated with various concentrations of HERL. After the treatment with HERL for 48 h, 30 µL of 3-(4,5-dimethylthiazol-20yl)-(3-carboxymethoxyphenyl)-2-(4-sulphophenyl)-2H-tetrazolium) (MTS) solution was added to each well and the cells were incubated for 2 h at 37℃. MTS solution was analyzed using a microplate reader (BioTeck Instruments, Winooski, VT, USA) at 490 and 690 nm (background).
FACS analysis for sub-G1 DNA measurement
After treatment with HERL for 48 h, the detached HN22 and SCC15 cells were collected and combined with the adherent cells that had been released by trypsinization. The cells were fixed in 70% ethanol overnight at -20℃. The cells were subsequently stained with 0.02 mg/mL PI. The data were acquired using a FACScan flow cytometer.
DAPI staining
The number of cells with nuclear condensation and fragmentation was measured using DAPI staining. After treatment with HERL or 0.1% NaCl (control), the HN22 and SCC15 cells were harvested by trypsinization. The cells were resuspended in PBS, deposited on poly-L-lysine-coated slides, stained with DAPI solution (2 µg/mL) and observed under a fluorescence microscope.
Western blot analysis
After treatment with HERL, HN22 and SCC15 cells were harvested. The protein supernatant fractions were subjected to SDS-PAGE and then transferred to polyvinylidene difluoride (PVDF) membranes and blocked with 5% skim milk followed by hybridization with the indicated antibodies. After incubation with the horseradish peroxidase (HRP)-conjugated secondary antibody, the immunoreactive protein bands were observed using a chemiluminescence detection kit.
Results
HERL decreases the cell viability in oral cancer cell lines
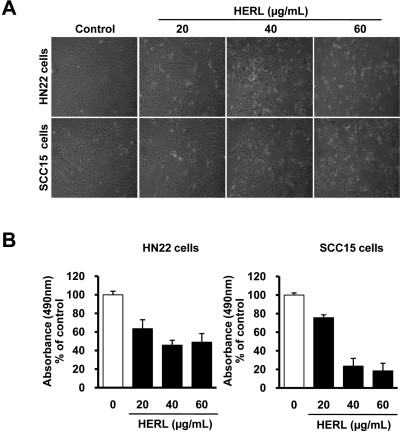
We first investigated the effect of HERL on the growth of HN22 and SCC15 cells. HERL-treated cells were detached from the dishes compared to 0.1% NaCl-treated cells and these morphological changes occurred in a dose-dependent manner (Figure 1A). We also measured the effect of HERL on cell viability in the MTS assay. The results showed dose-dependent decreases of cell viability in HN22 and SCC15 cells (Figure 1B). These results suggest that HERL has a growth-inhibitory effect on HN22 and SCC15 cells.
HERL induces apoptosis in oral cancer cell lines
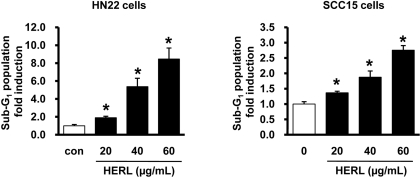
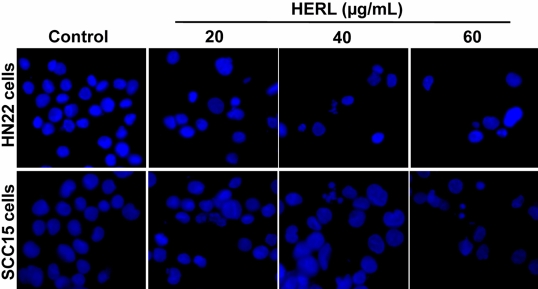
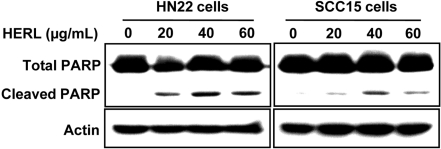
To examine whether HERL induced apoptosis, the number of cells accumulated in the sub-G1 phase of the cell cycle was counted by flow cytometric analysis. As shown in Figure 2, the sub-G1 population of HERL-treated cells accumulated in a dose-dependent manner compared to the vehicle control. To confirm the induction of apoptosis by HERL in HN22 and SCC15 cells, DAPI staining was performed and showed the fragmentation and condensation of nucleus in the cells treated with HERL (20, 40 and 60 µg/mL) for 48 h compared to the control (Figure 3). We next determined the level of PARP cleavage by Western blot analysis (Figure 4). After HERL treatment for 48 h, cleaved PARP was detected in the HN22 and SCC15 cells. Thus, we clearly showed that HERL induced apoptosis in HN22 and SCC15 cells to inhibit cell growth.
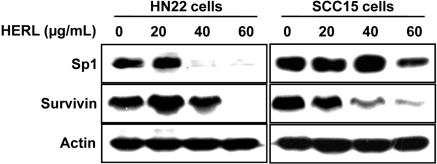
HERL inhibited anti-apoptotic protein Sp1 and survivin in oral cancer cell lines
In previous studies, it was reported that targeting of Sp1 protein is a good treatment strategy for various cancer cells (Lou et al, 2005; Safe and Abdelrahim, 2005; Abdelrahim et al, 2006; Papineni et al, 2009; Lu and Archer, 2010) and Sp1 protein regulates survivin protein as the downstream protein to induce apoptosis in cancer cells (Shin et al, 2010). Therefore, we assessed whether HERL affects Sp1 and survivin protein to induce apoptotic cell death in HN22 and SCC15 cells. As shown in Figure 5, HERL-treated HN22 and SCC15 cells exhibited decreased Sp1 and survivin expression in a dose-dependent manner. These results suggest that HERL might inhibit Sp1 and survivin protein to exert its apoptotic activity in HN22 and SCC15 cells.
Discussion
Natural extracts from edible plants have been identified as excellent candidates for cancer therapeutics based on safety and efficacy (Stewart et al, 2003). Recently, Rheum undulatum L. was screened for anti-cancer activity in vitro using human breast, ovary, cervix and lung cancer cell lines (Kang et al, 2008). In that study, emodin showed strong cytotoxicity in both estrogen receptor (ER)-positive and -negative breast cancer cell lines (Kang et al, 2008). In addition, HERL was shown to inhibit angiogenic activity in a zebrafish model (He et al, 2009). These studies suggest that HERL has potential benefits for the treatment of various cancers. However, the anticancer activity of HERL against oral cancer cells has not been well established. Therefore, we focused on the anticancer activity of HERL in human oral cancer cells. Our first purpose was to examine the anti-proliferative and pro-apoptotic effects of HERL on oral cancer cell lines and the other was to determine its molecular mechanism in HERL-induced apoptosis. First, we found that HERL decreased the number of viable cells in a dose-dependent manner, showing that HERL has an inhibitory effect on cell growth in HN22 and SCC15 cells. We found that HERL induced the apoptosis of HN22 and SCC15 cells as evidenced by the accumulation of sub-G1 phase cells, distinct chromatin condensation and nuclear fragmentation of the nucleus and the cleavage of PARP protein, a typical apoptotic marker.
Sp1 is a member of the mammalian transcription factor family that binds to GC-rich sites containing GC-boxes. Recently, it was reported that Sp1 protein is over-expressed in many human tumors and cancer cell lines (Zannetti et al, 2000; Chiefari et al, 2002; Wang et al, 2003; Hosoi et al, 2004; Yao et al, 2004). Sp1 has also been implicated in multiple cell processes, including apoptosis, through activation of the FAS-ligand (Kavurma et al, 2001; Sun et al, 2001). In addition, survivin, an inhibitor of apoptosis and a key regulator of mitosis, is up-regulated in a variety of cancer cells and is often associated with a worse prognosis (Sun et al, 2001). Interestingly, several studies have shown that survivin expression can be regulated by Sp1 protein because it contains highly GC-rich sequences within its promoters (Li and Altieri, 1999; Wu et al, 2005; Li et al, 2006; Shim et al, 2010). Therefore, we examined the effects of HERL on the expression of Sp1 and its downstream target, survivin, as the key molecular factors in human oral cancer cells. Our results show that HERL decreases the expression levels of Sp1 and survivin proteins, suggesting that HERL inhibits the growth and induces apoptosis in HN22 and SCC15 cells through the regulation of Sp1 and survivin.
In summary, our data showed HERL exerts pro-apoptotic and anti-proliferative effects in HN22 and SCC15 human oral cancer cells and that Sp1 and survivin play important signaling roles in HERL-induced apoptosis. Therefore, we suggest that HERL is a promising anti-cancer drug candidate for the effective treatment of oral cancer.
 XML Download
XML Download