

 PDF
PDF ePub
ePub Citation
Citation Print
Print


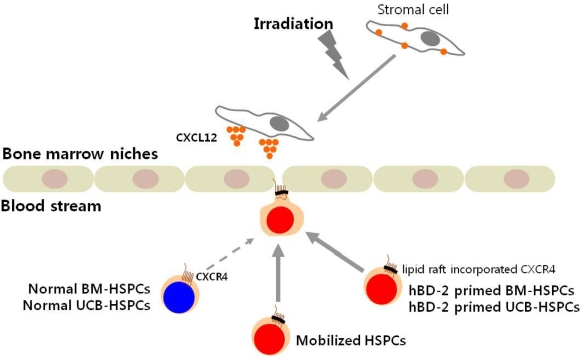
Hematopoietic stem/progenitor cells (HSPCs) circulate in peripheral blood (PB) under steady state conditions at very low levels to keep a pool of stem cells in balance in the bone marrow (BM) microenvironment [1-4]. Therefore, the bloodstream might be the "highway" by which HSPCs relocate between circulation in blood vessels and lodgment in BM endosteal and endothelial niches.
HSPCs can be mobilized from BM into PB during tissue injury [5,6], as well as after treatment of several mobilizing agents. Granulocyte colony-stimulating factor (G-CSF) is currently the most frequently used drug that may efficiently mobilize HSPCs after a few consecutive daily injections [7-9]. On the other hand, the collective term 'homing' refers to the early stages of BM seeding by HSPCs after transplantation, which precedes engraftment, proliferation, and differentiation of these cells into the hematopoietic lineage. In general, homing, engraftment, and expansion of HSPCs after transplantation are closely related processes [10,11].
Transplantation of HSPCs is dependent on the successful homing, engraftment, and repopulation of stem cells in the BM [12,13]. Stem cell homing through the circulatory system to the BM is the critical first step in this process.
The interaction of CXCL12 with its receptor CXCR4, which is expressed in HSPCs, has been implicated as a primary axis governing HSPC homing and engraftment into the BM following transplantation [14-16]. Current strategies to accelerate hematopoietic reconstitution after transplantation include transplantation of greater numbers of HSPCs or ex vivo expansion of harvested HSPCs before transplant. However, the number of cells available for transplantation is usually low, and strategies to expand HSPCs and maintain equivalent engraftment capability ex vivo are limited [17,18].
The migration of HSPCs toward a CXCL12 gradient has been correlated with the repopulating potential of transplanted HPSCs. Indeed, it has been demonstrated that successful homing and engraftment of HSPCs is significantly reduced if the CXCR4-CXCL12 axis is impeded [19].
Of note, it has been shown that leukapheresis products (i.e., C3 liquid-phase cleavage fragments, C3a) are known to enhance responsiveness of HSPCs to a CXCL12 gradient [20,21]. The molecular explanation of this phenomenon was that C3a increases CXCR4 incorporation into membrane lipid rafts, thus facilitating its more effective assembly with downstream signaling molecules [22].
In fact, C3a is one of the antimicrobial cationic peptides (AMPs), which have antimicrobial activity [23]. AMPs have been demonstrated to kill bacteria, enveloped viruses, fungi, and even transformed or cancerous cells without affecting the viability of eukaryotic cells. Selective effects of AMPs, like membrane perturbation of eukaryotic cells and killing of prokaryotic cells, are known to be dependent on the characteristics of membranes, such as the presence of cholesterol and membrane electric charge [24].
In addition, leukapheresis products which are enriched in activated granulocytes are known to release several cationic peptides and activate complement cascades [13]. We recently noted that activated granulocyte-derived cationic peptide positively primes responsiveness of murine HSPCs to a CXCL12 gradient [25].
Based on these data, we hypothesized that the use of cationic peptides as priming drugs might also enhance homing and engraftment of HSPCs after transplantation. In this study, we tested if human β defensin-2 (hBD-2), a well-known antimicrobial cationic peptide released from activated granulocytes, enhances the homing and engraftment of HSPCs after transplantation.
Materials and Methods
Animals
C57BL/6 mice were adopted at 1-2 weeks and used for experiments at age 6 to 8 weeks. Animal studies were approved by the Animal Care and Use Committee of the Institute of Laboratory Animal Resources, Seoul National University.
Bone marrow nucleated cells (BMNCs)
BMNCs were prepared by flushing femurs and tibias of pathogen-free, 6- to 8-week-old mice without enzymatic digestion. They were lysed with BD Pharm Lyse buffer (BD Biosciences, San Jose, CA, US) to remove red blood cells (RBCs), washed, and resuspended in appropriate media for further analysis.
Trans-well migration assay
The trans-well migration assay was performed as described elsewhere [25]. Briefly, unless otherwise indicated, RBC-lysed BMNCs from C57BL/6 mice were resuspended in assay media (RPMI containing 0.5% bovine serum albumin) and equilibrated for 30 min at 37℃. Six hundred-fifty microliters of assay media in the presence or absence of CXCL12 (50-300 ng/mL, PeproTech, Rocky Hill, NJ, USA) was added to the lower chambers of a Trans-well 24-well plate (Costar Corning, Cambridge, MA, USA). hBD-2 was purchased from Prospec (East Brunswick, NJ, USA). Aliquots of normal or 30 min hBD-2-exposed cell suspension (1×106 cells/100 µL) were loaded onto the upper chambers with 5 µm-pore filters, which were incubated for 3 hours at 37℃, 95% humidity, and 5% CO2. Cells from the lower chambers were scored using FACS analysis for migration of BMNCs to test reagents. The results are presented as a migration index, i.e., fold increase of cell numbers migrated toward the lower well compared to control media. In some experiments, migrated cells harvested from the lower chambers were plated in semisolid clonogenic assays supplemented with cytokines and were stimulated to grow colony-forming units-granulocyte-macrophage (CFU-GM) and colony-forming units-macrophage (CFU-M) as described elsewhere [25].
Colony forming unit assay
Red blood cells were lysed with BD Pharm Lyse buffer. Nucleated cells were subsequently washed twice and used for CFU-GM and/or CFU-M colonies. BMNCs were resuspended in human methylcellulose base media provided by the manufacturer (R&D Systems, Minneapolis, MN, USA) supplemented with 25 ng/mL recombinant murine granulocyte macrophage colony-stimulating factor (mGM-CSF) and 10 ng/mL recombinant murine interleukin-3 (mIL-3) for CFU-GM, 10 ng/mL recombinant murine macrophage stimulating factor (mM-CSF) for CFU-M. Cultures were incubated for 7 days, at which time they were scored for the number of CFU-GM colonies under an inverted microscope.
Phosphorylation of intracellular pathway proteins
Western blot analysis was performed on extracts prepared from mouse EL4 cell lines that had been kept in RPMI medium containing 0.5% bovine serum albumin (BSA) to render the cells quiescent. Cells were stimulated with either a low or high concentration of CXCL12 (50 ng/mL or 300 ng/mL) in the presence or absence of hBD-2 for 90 sec at 37℃ and then lysed for 10 min on ice in M-Per lysing buffer (Pierce, Rockford, IL, USA) containing protease and phosphatase inhibitors (Sigma, St. Louis, MO, USA). Extracted proteins were separated on a 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gel and then transferred to a nitrocellulose membrane (Hybond ECL; Amersham Life Sciences, Little Chalfont, United Kingdom). Phosphorylation of 44/42 MAPK protein was detected by protein immunoblotting using mouse monoclonal 44/42 phospho-specific MAPK antibody (New England Biolabs, Beverly, MA, USA), with horseradish peroxidase conjugated goat anti-mouse immunoglobulin G (Santa Cruz Biotechnology, Santa Cruz, CA, USA) as secondary antibody. Equal loading in the lanes was evaluated by stripping the blots and reprobing them with the appropriate monoclonal or polyclonal antibodies. The membranes were developed with an ECL reagent (Amersham Life Sciences), dried, and exposed to film (HyperFilm; Amersham Life Sciences).
In vitro adhesion assay
Wild-type BM stromal cells were grown to confluence, and normal or 30 min hBD-2-exposed BMNCs were subsequently plated over BM-derived stroma feeder layers established in well plates. Five minutes later, plates were gently shaken, and non-adherent cells were discarded. Subsequently, the cultures were trypsinized and stimulated to grow CFU-GM colonies as mentioned earlier. The number of colonies was scored under an inverted microscope.
Evaluation of in vivo engraftment of transplanted HSPCs
To test whether hBD-2 priming enhanced homing and engraftment of transplanted HSPCs, we used a short-homing assay to evaluate lodging of HSPCs to the spleen and bone marrow. Normal recipient mice were lethally irradiated (950 cGy) and 24 hours later were transplanted with 5×105 normal or 30 min hBD-2-exposed BMNCs by tail vein injection. At 18 hours or 12 days after BMNC transplantation, spleens were removed and fixed for colony forming units-spleen (CFU-S) count. At the same time, BM were harvested from the femurs and plated in methylcellulose cultures and stimulated to grow CFU-GM colonies. CFU-S was also counted.
Results
CXCL12 directly chemoattracts BM cells in a dose-dependent manner
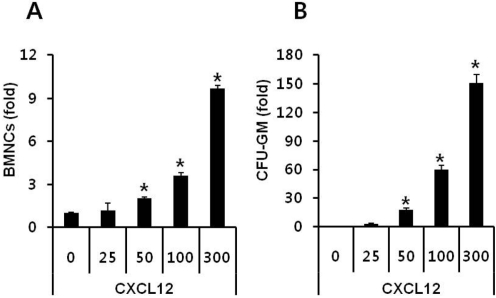
To optimize the trans-well migration assay we tested different doses of CXCL12 using a 5 µm pore sized trans-well. CXCL12 directly chemoattracted not only BMNCs (Figure 1A), but also CFU-GM progenitor cells (Figure 1B) in a dose-dependent manner. However, CXCL12 at a concentration lower than 25 ng/mL did not chemoattract either BMNCs or CFU-GM progentitor cells efficiently. Significant migrations of BMNCs to CXCL12 were shown in concentrations of 50 ng/mL or higher. Although 50 ng/mL of CXCL12 is relatively high compared to its physiological plasma concentration [26], we chose 50 ng/mL of CXCL12 as the standard concentration for our experiments based on the results obtained from our optimization assay.
hBD-2 stimulation enhanced responsiveness of HSPCs to a CXCL12 gradient in a dose-dependent manner
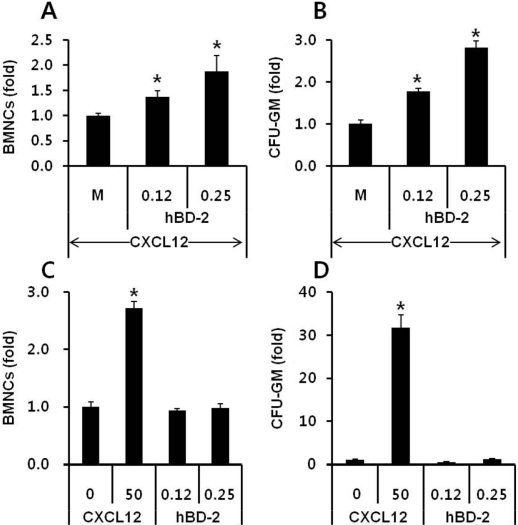
The antimicrobial cationic peptide hBD-2, which is known to be released from activated human granulocytes, enhanced responsiveness of BMNCs to a CXCL12 gradient. As expected, the migration of hBD-2-exposed BMNCs to a CXCL12 gradient was significantly enhanced compared to that of normal BMNCs (Figures 2A and 2B). This accelerated migration of BMNCs and CFU-GM progenitor cells to a CXCL12 gradient was dependent on the hBD-2 concentration used. However, hBD-2 itself did not show a direct chemotactic effect on BMNCs (Figure 2C) and CFU-GM progenitor cells (Figure 2D). Considering hBD-2 is known to use CCR6 for signal transduction, the hBD-2 priming effect was not a receptor-mediated phenomenon. MIP-3α, which is known as an agonist for CCR6, did not enhance migration of BMNCs to the CXCL12 gradient (data not shown). These results strongly support that enhanced migration of hBD-2-primed cells might be mediated by enhancing incorporation of CXCR4 into lipid rafts as similar to what was shown in C3a primed cells [20,21].
hBD-2 enhanced CXCL12-CXCR4-mediated intracellular signaling strength
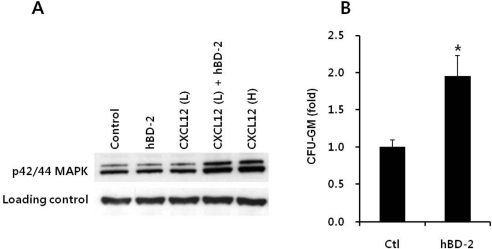
We tested if enhanced CXCR4 incorporation into membrane lipid rafts in the presence of hBD-2 is better connected to downstream signaling machinery and can respond robustly to a CXCL12 gradient, as a previous report showed that the supernatants of leukapheresis products increased p42/44 MAPK phosphorylation mediated by enhancing incorporation of CXCR4 into membrane lipid rafts [20,21].
We noticed that hBD-2 itself does not induce phosphorylation of p42/44 MAPK but enhanced p42/44 MAPK phosphorylation induced by CXCL12 in EL4 cells. CXCL12-induced phosphorylation of p42/44 MAPK was strongly enhanced in hBD-2-primed cells (Figure 3A). This also strongly supports that the priming effect of hBD-2 is not mediated by receptor-ligand interactions.
hBD-2 priming increased the adhesion of CFU-GM progenitor cells to BM stromal cells
We questioned the effects of priming agents on other processes essential for the engraftment of HSPCs, as they must adhere to the BM stroma to proliferate and differentiate. To address this question further, we carried out in vitro adhesion experiments with BMNCs. We found that hBD-2-exposed CFU-GM progenitor cells showed increased adhesion (Figure 3B). We suggest that hBD-2, by enhancing responsiveness of HSPCs to a CXCL12 gradient, may directly modulate HSPC homing.
hBD-2 exposure to BMNCs accelerated homing/engraftment after transplantation into lethally irradiated mice
Based on our in vitro results showing enhanced migration and adhesion, we examined whether hBD-2-primed BMNCs engraft faster than normal BMNCs. As summarized in Table 1, hBD-2-primed BMNCs homed better to the BM and spleen of lethally irradiated recipient mice. CFU-GM progenitor cells at 18 hours after BMNC transplantation were increased more than two fold in the femur of hBD-2-primed BMNC transplanted group compared to the normal BMNC transplanted group (data not shown). No CFU-GM or CFU-S was recovered from lethally irradiated control mice that did not receive BMNCs.
hBD-2 exposure to BMNCs did not affect the viability and differentiation potential of BM progenitor cells
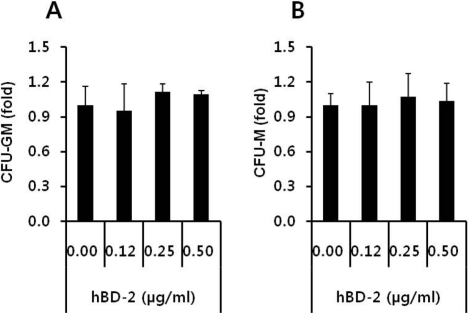
Finally, we evaluated the safety of hBD-2 exposure on hematopoietic progenitor cells. BMNCs (5×105/mL) were incubated for 60 min at 37 CO2 incubator in the absence or presence of hBD-2 at 0.12, 0.25, or 0.50 µg/mL concentrations. These cells were subsequently examined via in vitro colony forming assays for the number of CFU-GM and CFU-M progenitor cells. As expected, hBD-2 exposure did not affect the viability and the clonogeneic potential of HSPCs (Figures 4A and 4B).
Discussion
Homing is the first step into BM seeding by HSPCs after transplantation, which precedes their engraftment, proliferation, and differentiation. It is also well established that the CXCL12-CXCR4 axis plays a pivotal role in directing the homing of HSPCs from the PB to the BM [10,27,28]. While CXCL12 is expressed in the BM stroma, CXCR4 is present on the surface of HSPCs [29,30]. It is not surprising that proper functioning of the CXCL12-CXCR4 axis is crucial in directing the homing and engraftment of HSPCs into the BM after transplantation.
It has been recently demonstrated that CXCR4 requires an association with membrane lipid rafts for proper signaling function [31-34]. Additionally, it has been shown that membrane lipid rafts are important for T-cell polarization and chemotaxis [35]. Increased responsiveness of HSPCs to a CXCL12 gradient in the presence of other factors was named as the priming effect [36]. Previous work revealed that several small molecules (i.e., fibronectin and fibrinogen fragments, hyaluronic acid, granulocyte-derived cationic peptides, and C3a) that are present in supernatants from leukapheresis products may modulate the responsiveness of HSPCs to a CXCL12 gradient by enhancing the incorporation of CXCR4 into membrane lipid rafts [37-39].
In this study, we evaluated if the cationic peptide hBD-2 released from activated granulocytes will enhance homing and engraftment of transplanted HSPCs. We found that HSPCs stimulated by hBD-2 better responded to a CXCL12 gradient. Moreover, we found for the first time that transplantation of hBD-2-primed BMNCs into lethally irradiated recipients results in an increase in the number of donor-derived CFU-granulocyte-macrophage progenitors in the bone marrow cavities, as well as an increase in formation of CFU-S.
hBD-2 itself did not show chemotactic activity but an enhanced migration of HSPCs to a CXCL12 gradient compared to normal cells. Pretreatment of HSPCs with hBD-2 enhanced CXCL12-CXCR4-mediated intracellular signaling, adhesion to stromal cells, and their engraftment into irradiated mice, further emphasizing that hBD-2 plays an essential role in promoting the homing and engraftment of HSPCs to the bone marrow through its synergistic interaction with the CXCL12-CXCR4 signaling axis.
Considering the fact that CXCL12, secreted by bone marrow stromal cells, regulates the migration of HSPCs and thereby plays a central role in anchoring stem cells to the bone marrow microenvironment after transplantation, the sensitization of the CXCL12-CXCR4 axis by hBD-2 could be used to accelerate the homing and migration of transplanted HSPCs into the BM.
Activated granulocyte-derived AMPs might be safer than previously described priming agents (C3a, angiopoietin etc.) [20,21,40], because AMPs have been demonstrated to kill bacteria, enveloped viruses, fungi, and even transformed or cancerous cells without affecting the viability of eukaryotic cells. Selective effects of AMPs, like membrane perturbation of eukaryotic cells and killing of prokaryotic cells, are known to be dependent on the characteristics of membranes, such as cholesterol content or membrane electric charge. In agreement with our hypothesis, hBD-2 exposure did not affect the viability and differentiation potential of HSPCs.
Taken together, we suggest that hBD-2 might be employed as a new agent to prime ex vivo HSPCs before transplantation. This novel approach would be particularly important in umbilical cord blood transplantation, where the number of HSPCs available for reconstitution of recipient is often limited.

 XML Download
XML Download