

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Buckwheat is one of favorite foods in Asian countries. Buckwheat is a potent major allergic food, especially the most important causes of allergy among grains commonly consumed in Korea.12 Moreover, it is one of leading causes of anaphylaxis.3 The consumption of buckwheat in Western countries has also increased recently due to its gluten-free characteristics,4 and it has been described as a hidden allergen in Italian cuisine, such as pizza and pasta.5
Labeling of allergens in food merchandise is crucial, and most developed countries legally require labeling of common food allergens. Laws for the labeling vary among countries, despite the recommendations by the Codex Alimentarius Commission.6 Buckwheat is a major cause of food allergy in Korea, and Korean Food and Drug Administration (FDA) has designated labeling of buckwheat in foods as mandatory (KFDA official announcement 2015-77). However, labeling does not provide detailed information on the content of specific allergens, and it would be helpful to know the concentration of these allergenic molecules in food products.
Various 9-, 16-, 19-, and 24-kDa protein allergens have been described as the major allergens from buckwheat.7 A 19-kDa allergen homologous to vicilin was more specifically recognized by allergic patients than by asymptomatic subjects. Specific immunoglobulin E (IgE) measurements against the recombinant 19-kDa allergen had better diagnostic value than those of whole buckwheat extracts.89 The 19-kDa allergen was designated as Fag e 3, according to the guidelines of the International Union of Immunological Societies Allergen Nomenclature Subcommittee.
Quantification of major allergen in extracts may be essential for standardization of allergen, pathogensis, and epidemiologic studies. Various methods have been used for the quantification of major allergens, such as radial immunodiffusion using polyclonal antibodies or 2-site enzyme-linked immunosorbent assay (ELISA) kit. Mass spectrometry techniques could be also applied to detect and quantify the allergens in foods.10
In this study, we produced monoclonal antibodies (mAbs) to recombinant Fag e 3 and developed a method to quantify Fag e 3 allergen concentrations.
MATERIALS AND METHODS
Allergen extracts
Buckwheat and walnut extracts were prepared as described previously.7 In brief, buckwheat flour was extracted with phosphate buffered saline for 72 hours at 4℃ after defatting with ethyl ether. The supernatant was dialyzed against distilled water for 48 hours after centrifugation at 50,000 g for one hour at 4℃. Subsequently, the extract was lyophilized and stored at -20℃ until use. The protein concentration of the reconstituted extract in phosphate buffered saline was determined by Bradford assay (Bio-Rad, Hercules, CA, USA). Peanut extract was kindly provided by Dr. Kyung Eun Lee (Department of Pediatrics, Yonsei University College of Medicine).
mAbs to recombinant Fag e 3
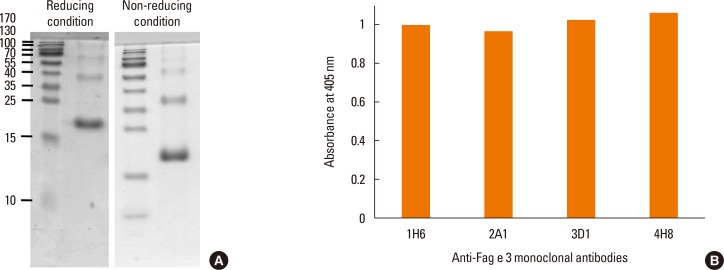
Recombinant Fag e 3 was expressed in Escherichia coli and purified using a Ni-column from inclusion bodies, as described previously.8 Mouse mAbs to Fag e 3 were produced by the fusion of myeloma cells (Sp 2.0-Ag 14) and spleen cells from BALB/c mice immunized with recombinant Fag e 3 3 times at 2-week intervals. The hybridomas that produced antibodies against recombinant Fag e 3 were screened by ELISA and cloned by limiting dilution. mAbs from the expanded clones were purified with protein G (Sigma-Aldrich, St. Louis, MO, USA). All of the antibodies were IgG1 according to mouse mAb isotyping reagents (Sigma-Aldrich).
Sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) and immunoblot analysis
Recombinant protein or buckwheat extract was run on a 15% acrylamide gel containing SDS under reducing or non-reducing conditions. The gel was stained with Coomassie blue or transferred to a polyvinylidine difluoride membrane (0.45 µm; Millipore, Bedford, MA, USA). The membrane was blocked overnight with 3% skim milk in Tris-buffered saline containing 0.05% Tween 20. Subsequently, membranes were reacted with hybridoma culture supernatant and then incubated with 1:1,000-diluted goat anti-mouse IgE conjugated with alkaline phosphatase (Sigma-Aldrich). Color was developed with 3-bromo-4-chloro-5-indolyl-phosphate, and nitro blue tetrazolium as a substrate (Promega, Madison, WI, USA).
Biotinylation of antibody
mAbs purified with a protein G column were biotinylated with EZ-Link® Sulfo-NHS-LC-Biotin (Thermo Fisher Scientific, Waltham, MA, USA). In brief, 2 mg/mL of antibodies were incubated with NHS-LC-Biotin on ice for 4 hours, and unreacted NHS-LC-Biotin was removed by extensive dialysis against phosphate buffered saline.
Measurement of Fag e 3 using a 2-site ELISA
The optimal antibody combination was established by comparing titration curves of recombinant Fag e 3 using the 3 different antibodies. The combination of 3D1 (10 µg/mL) as a capture antibody and biotinylated 4H8 (1:1,000-diluted) as a detection antibody gave optimal results. Antigens were diluted in buffers containing 1% of various detergents (Triton X, Nonidet P40, or SDS) to facilitate the dissociation of Fag e 3 in the extract. Some of the each sample was heated at 100℃ for 5 minutes after the sample was diluted in detergent buffer, and compared the results with samples without heat treatment.
In brief, the capture antibody was diluted in carbonate buffer (pH 9.6) and was coated overnight at 4℃. Serially diluted antigens (recombinant Fag e 3 or allergen extracts) were added to the wells and incubated for one hour at room temperature after blocking with 3% skim milk. This was then incubated for one more hour after adding the detection antibody, and incubated again with streptavidin-conjugated peroxidase (Sigma-Aldrich) for 30 minutes. Microtiter plates were washed at least 3 times with phosphate-buffered saline containing 0.05% Tween 20 between each step. Color was developed with 2,2′-azino-bis (3-ethylbenzothiazoline-6-suphonic acid) (Thermo Fisher Scientific) or 3.3′, 5,5′-tetramethylbenzidine (Kirkegaard Perry Laboratories, Gaithersburg, MD, USA).
For comparison, the concentration of Ara h 1, a vicilin-like allergen from the peanut, was also determined with the Ara h 1 ELISA kit (Indoor Biotechnologies Inc., Charlottesville, VA, USA).
RESULTS
Specificity of mAbs
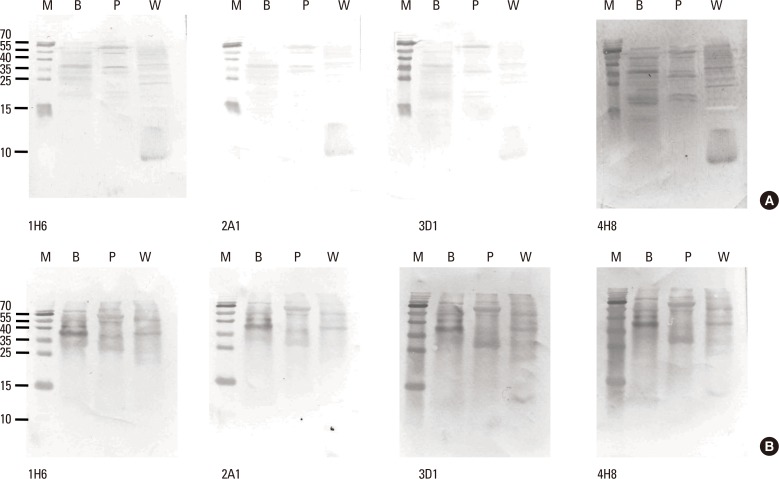
Reactivity of the mAbs to total buckwheat extract containing native allergens was examined by immunoblotting with mAbs, which were produced against recombinant protein. In the extract, Fag e 3 separated under reducing conditions was barely recognized by mAbs, despite mAbs showing good reactivity to the extract on ELISA (Fig. 1). This observation motivated us to perform the immunoblot again, while separating the extract under non-reducing conditions. Interestingly, all of the mAbs preferentially recognized a 38-kDa protein, putative dimerized Fag e 3. We also evaluated whether the mAbs could detect vicilin family major allergens of peanut (Ara h 1) and walnut (Jug r 2). Putative Ara h 1 (64 kDa) and Jug r 2 (44 kDa) under nonreducing condition only were also recognized by the all mAbs, respectively (Fig. 2).
Sensitivity of 2-site ELISA
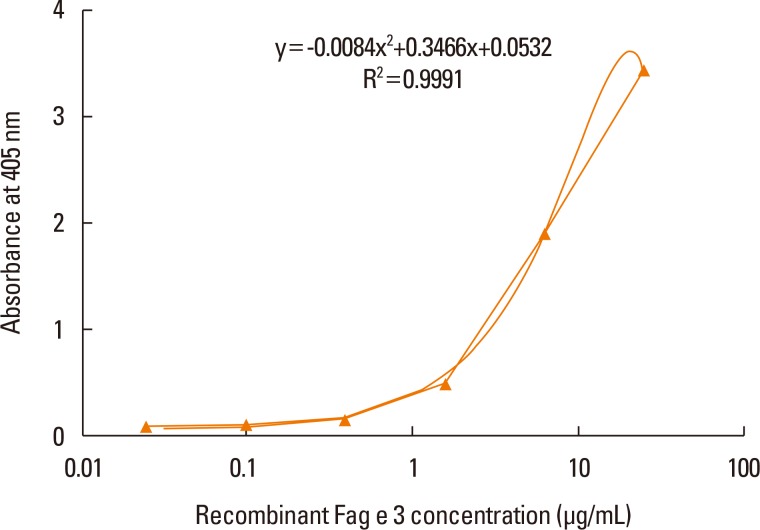
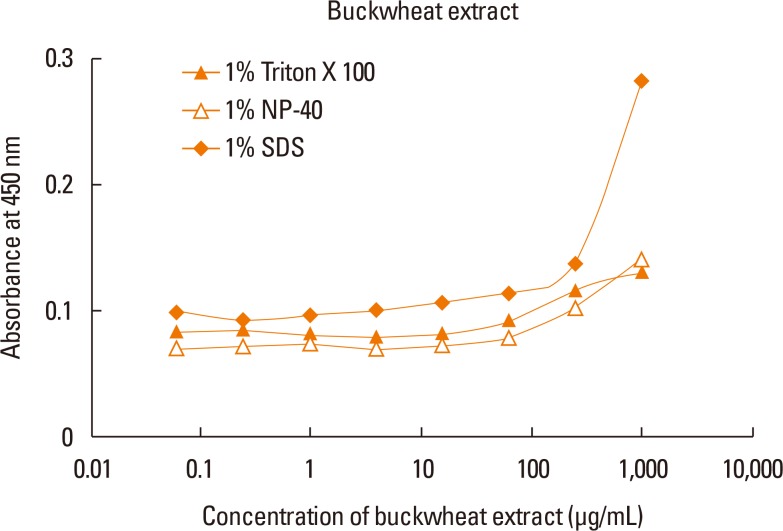
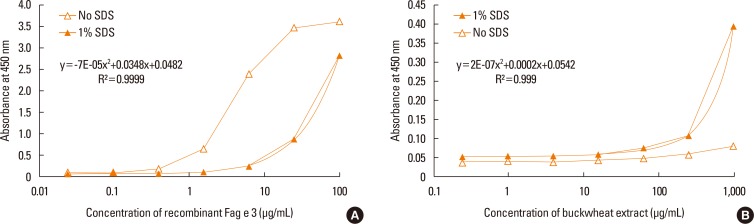
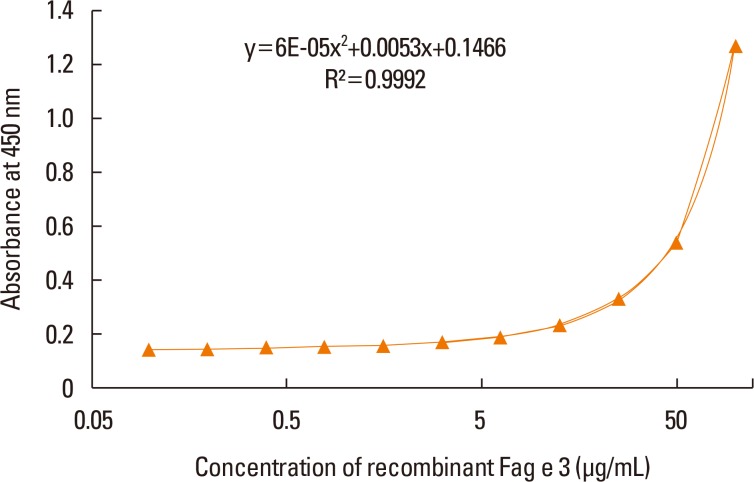
ELISA with selected antibodies was able to detect less than 1 µg/mL of recombinant Fag e 3 (Fig. 3). However, it could not detect native Fag e 3 in the total extract of buckwheat. Therefore, we added various detergents to facilitate the dissociation of Fag e 3 in the total extract (Fig. 4). An ELISA with SDS showed better detection, but had lowered overall detection capability. The 2-site ELISA system developed in this study was able to detect up to 6 µg/mL of the vicilin allergen when tested with recombinant Fag e 3. More than 100 µg/mL of total buckwheat protein is needed for efficient detection of Fag e 3 (Fig. 5).
However, the sensitivity of ELISA was improved by heating the antigen mixture at 100℃ for 5 minutes after the addition of 1% SDS. The detection limit of the 2-site ELISA was 0.8 µg/mL of recombinant Fag e 3 (Fig. 6). The detection limit of total buckwheat extract was also lowered to 60 µg/mL by heating the samples with the detergent. Vicilin-like allergens were not detected from peanut and walnut total extracts by the developed 2-site ELISA.
DISCUSSION
Immunoassays for the quantification of allergens in food are useful for allergen avoidance and standardization of allergen extracts.11 We have developed a 2-site ELISA to quantify the buckwheat allergen Fag e 3 using mAbs raised against recombinant Fag e 3.
All mAbs showed good reactivity to total buckwheat extract. However, they were not able to readily detect native Fag e 3 in the extract under reducing condition. All the mAbs produced against recombinant Fag e 3 seem to recognize the conformational epitopes of vicilin-like allergens. Ara h 1 (64 kDa) and Jug r 2 (44 kDa) were also recognized by the mAbs under non-reducing condition.
Interestingly, individual mAbs showed good response to the extract in ELISA experiment, suggesting good affinity to native Fag e 3. However, native Fag e 3 in the extract was not detected by the combination of 2 mAbs, although recombinant Fag e 3 was well quantified. mAbs to recombinant Fag e 3 may recognize epitopes very close to each other. Binding of one mAb to an epitope region may decrease the accessibility of another mAb to a close epitope, possibly due to steric hindrance. Partial denaturation of Fag e 3 by detergents may increase the accessibility of another mAb. Epitope mapping should be performed to explain this hypothesis. In addition, the binding of mAbs to Fag e 3 may be hampered by other proteins which associated with vicilins or by interactions between vicilin molecules, as they are known to form multimers. Typically, vicilins are disk-shaped trimeric proteins with various subunit compositions due to post-translational proteolytic processing and glycosylation.12 Glycosylation may also influence antibody recognition.13 IgE reactivity to native Jug r 2 without clinical significance was described due to cross-reactive carbohydrate epitopes.14 Structural similarity of vicilin-like proteins from peanut (Ara h 1), walnut (Jug r 1), hazelnut (Cor a 11), and cashew nut (Ana o 1) was described, indicating its potential cross-reactivity,15 and all mAbs produced by our team also have affinity to Ara h 1 and Jug r 1 under non-reducing condition.
Recombinant Fag e 3 had good IgE reactivity, indicating that it closely resembles its native counterpart.8 We did not observe structural differences in recombinant Fag e 3 based on reducing and non-reducing gels. However, reducing conditions may cause native Fag e 3 to undergo a slight change in its configuration, as it showed different reactivity to mAbs under those conditions. However, the structurally change under reducing conditions is believed to retain most of its IgE epitopes, as it displayed good IgE reactivity based on IgE immunoblot analysis.7 Thermal treatment (30-50 minutes) in the presence of polysaccharides can cause structural change to vicilin-like allergens. This leads to markedly reduced immunogenicity through the Maillard reaction. However Fag t 3, a vicilin-like allergen from tartary buckwheat (Fagopyrum tataricum), was stable even after incubation at 90℃ for 30 minutes.16 Therefore, we suspect that the Maillard reaction did not occur in this study during heat treatment with 1% SDS for 5 minutes at 100℃.
We used the detergent SDS to improve the detection of Fag e 3 in the dilution buffer. Approximately 12% of buckwheat protein was found to be vicilin. Detecting allergenic components from food that is mixed with various seasonings, additives, and other foods can be challenging. In the case of the peanut, improved detection of Ara h 1 by 2-site ELISA were obtained by heating the sample and adding nonfat dry milk to the extraction buffer (up to 400-fold).17 Therefore, heating the antigens and using detergent prior to serial dilution could improve the extraction and detection of buckwheat major allergen from various foods and environments. The sensitivity of our 2-site ELISA improved approximately 7.5 times by heating before serially diluting the antigens.
Ara h 1 is a well-characterized vicilin allergen. It is a heat stable protein with a stable trimeric form (64 kDa) that is mediated through hydrophobic interactions. This protein comprises 12%-16% of the total protein in peanut extracts.181920 Ara h 1 was measured not only in peanut and food extracts, but also in various environments.172021 Buckwheat could be sensitized by inhalation in occupational and domestic exposure, mainly due to buckwheat chaff-stuffed pillows contaminated with buckwheat flour,22 or buckwheat flour in buckwheat noodle factory.23 It is likely that Fag e 3 can be measured in various environmental dusts such as restaurants that serve buckwheat, factories that produce buckwheat noodles, in addition to meals that contain buckwheat.
The mAbs produced against recombinant Fag e 3 also recognize vicilin-like allergens from tree nuts. Vicilin-like allergens are also described in black walnut (Jug n 2), lentil (Len c 1), pea (Pis s 1), cashew (Ana o 1), hazel (Cor a 1), and sesame (Ses i 3). Clinically relevant cross-reactivity between the pea and the peanut was reported due to vicilin-homologous allergens, Pis s 1, and Ara h 1.24 However, there is a low rate of clinical cross-reactivity among these vicilin belonged allergens.
Buckwheat 2-site ELISA kits are already produced by Crystal Chem Inc. (Downers Grove, IL, USA) and ELISA systems (Windsor, Queensland, Australia). These kits utilize buckwheat extract for the surveillance of buckwheat allergen contamination in foods. The food industry and official food control agency laboratories in Japan employ the ALLERGENEYE ELISA kit (Justia Legal Resources, Tokyo, Japan), which detects a 24-kDa allergen (a putative Fag e 1), and the FASTKIT ELISA (COSMO Bio Co., Ltd., Tokyo, Japan) (Notification No. 1106001, 2002).25 A buckwheat DNA detection system (Shimadzu Cor., Kyoto, Japan) is also available. Employing better detection systems for buckwheat allergens to complement the current food allergy labeling system is an important goal. The detection of clinically relevant allergens such as Fag e 2 or Fag e 3 shows better diagnostic value than that of the total extract.826 Furthermore, the importance of prevention of food allergy is being emphasized in a public health problem.27
We hope that the Fag e 3 ELISA developed in this study is helpful for the standardization of buckwheat extract as well as the detection of buckwheat allergens in foods and environments.
 XML Download
XML Download