

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Chronic rhinosinusitis (CRS) is one of the common diseases affecting about 10% of the general population, causing significant socioeconomic burden.12 The phenotypes of CRS, clinically observable characteristics, have been classified based on endoscopic or computed tomographic findings of nasal polyps (NP),3 the presence of lower airway disease (e.g. aspirin-exacerbated respiratory disease), and hypersensitivity to allergens (e.g. allergic fungal rhinosinusitis). Furthermore, CRS can also be subdivided by underlying diseases, such as cystic fibrosis, ciliary dyskinesia, autoimmune diseases, immune deficiency, and systemic illness.456 Specifically, NP are presented as a severe form of inflammation and remodeling among the phenotypes of CRS, and the presence of NP has been considered the most important clinical aspect of phenotyping CRS due to therapeutic challenges. For this reason, histologic subtypes of NP have been sought to be defined more precisely: eosinophilic or non-eosinophilic NP. Mucosal eosinophilia predicts recurrence after surgeries and risks of having comorbidities.7 However, there are no standard methods to evaluate tissue eosinophilia due to uneven distribution throughout the tissue.8 Multiple inflammatory cells exist in most NP tissue, but the function and pathological roles of these cells are not yet completely elucidated.9 Even in eosinophilic NP, neutrophils and macrophages exist and may play a role in nasal polypogenesis.2 Therefore, the subtyping of CRS or NP based on inflammatory cells has limitations. One phenotype can include several molecular mechanisms (endotypes), and one endotype can also involve several phenotypes. Due to these challenges, it is hard to design a personalized treatment approach with clinical phenotyping or histologic subtyping alone.
The concept of personalized medicine in the field of CRS can be incorporated after establishing endotypes. This is because CRS involves various pathomechanisms showing high heterogeneity, which causes different therapeutic responses. Endotyping also helps determine optimal primary therapeutic modality, select a good responder to a specific treatment, and predict treatment outcomes and risks for comorbidities, such as asthma.
We herein summarize pathophysiology and potential endotypes of CRS with NP (CRSwNP) and introduce biologicals available now or under investigation in CRS.
PATHOGENESIS AND ENDOTYPES OF CRS
Epithelial barrier function and epithelial cytokines
CRS can be understood as a dysfunctional host-environment interaction at the nasal and sinus mucosa.1011 Bacterial and viral infections, fungal extracts, and protease allergens play potential roles in CRS as external stimuli.12 For the evidence of bacterial roles, the superantigen hypothesis has been strongly suggested. Colonized Staphylococcus aureus secretes enterotoxins that augment local T helper 2 (Th2) inflammation and may play a role in the pathogenesis of NP as a disease modifier rather than an initiator.13 Another hypothesis of bacterial roles is that bacterial biofilms may be involved in CRS pathogenesis. Biofilms are organized structures comprised of bacteria protectively encased within extracellular matrix (ECM), which is associated with local inflammatory markers and disease severity.14 Although many studies have attempted to find evidence, the role of fungi in CRS has drawn lots of controversies in the last decade.15 There is still relatively sparse evidence that viral infection is a chronic source of CRS or involved in the initiation and development of sinonasal allergic inflammation. However, ds-RNA, a toll-like receptor 3 (TLR3) ligand, is a potent inducer of thymic stromal lymphopoietin (TSLP)16 and a potential changer of innate lymphoid cell (ILC) phenotypes,17 which may be associated with chronicity of airway disease. Moreover, viral infection has been clearly implicated in the exacerbation of allergic airway diseases, such as asthma, in a number of studies.18
The epithelial barrier is the first line of defense; its breakdown can play a significant role in allowing external stimuli to enter nasal tissue and provoke immune responses.19 Functional and mechanical defects have been reported in NP. The protease-activated receptor (PAR) contributes to the production of cytokines and chemokines from the epithelium in response to external stimuli, such as bacteria, fungi, and allergens.1220 Epithelial barrier destroyed by protease activities enables allergens to pass physical epithelial barriers, culminating in allergen sensitization.21 It also signals epithelial cells to secrete innate cytokines and then facilitates the induction of eosinophilic inflammation. Epithelial-derived innate cytokines, such as interleukin (IL)-25, IL-33, and TSLP, may also participate in the evolution of NP.22 IL-33 is secreted by immune cells, such as macrophages and dendritic cells as well as epithelial cells.23 Full-length IL-33 is extracellularly released when epithelial cells undergo necrosis and necroptosis via tissue damage caused by external stimuli. Biologically active full-length IL-33 plays a role in mucosal inflammation recruiting neutrophils via chemokines, including chemokine (C-X-C motif) ligand (CXCL)-1 and CXCL-2.242526 Gordon et al.27 have suggested a potential role of IL-33 in eosinophilic inflammation by demonstrating a splice variant of IL-33 missing exons 3 and 4, which localizes to the cytoplasm of epithelial cells, is actively released and strongly related to Th2 inflammation, whereas full length is not. Several studies have sought to investigate the expression and role of IL-33 in CRS. There have been conflicting results of the expression of IL-33 in CRS. It has been reported that IL-33 mRNA is highly expressed in nasal mucosa but was not elevated in NP or other inflamed areas of the sinuses in CRSwNP.282930 A significant up-regulation of ST2 expression has been demonstrated in ethmoid mucosa from CRSwNP, but the concentration of IL-33 protein is not significantly different between nasal polyp and control tissue.31 Kim et al.23 have recently demonstrated that IL-33 is up-regulated in other CRS tissues compared to eosinophilic NP and correlates with Th1/Th17 cytokines. IL-33 may contribute to the induction of different types of inflammation under various microenvironments.
IL-17E, also known as IL-25, is released by Th2 cells, mast cells, eosinophils as well as epithelial cells. It is produced and stored in the cytoplasm of epithelial cells as a result of external stimuli, including allergen proteases.32 IL-25 transcript levels have been reported to increase in CRS tissues, including NP and to correlate with disease severity and blood eosinophilia,3033 whereas an earlier study has reported that IL-25 and GATA-binding protein 3 (GATA-3) transcripts were decreased in NP vs control tissues.29 Additionally, polyp-derived IL-17RB (+) Th2 cells were identified in NP, which co-expressed ST2 and enhanced IL-5 and IL-13 production in response to IL-25 and IL-33.34 Protein levels of IL-25 are up-regulated in non-eosinophilic NP as well as eosinophilic NP.2235 Of note, the fact that IL-25, known as a cytokine involved in diverse Th2-mediated diseases, also correlated with inflammatory mediators involved in Th1 and Th17 responses in Asian subjects suggests that it may play diverse roles in polypogenesis besides promoting Th2 inflammation.3336 Blockade of IL-25 reduced the burden of NP in a mouse model of NP and represented a potential novel therapeutic target.36
TSLP is well known to be induced in airway epithelial cells by viruses, TLR3 agonists, protease, and pro-inflammatory cytokines.16373839 IL-1β and tumor necrosis factor (TNF)-α regulate TSLP transcript expression in an nuclear factor-kappa B (NF-κB)-dependent manner.39 Several researchers have demonstrated that TSLP mRNA is overexpressed in eosinophilic NP and associated with Th2 inflammation.404142 TSLP induces the differentiation of naïve T cells into effector Th2 cells via enhancement of OX40 ligand (OX40L)-OX40 axis on the interaction between dendritic cells and CD4 T cells.43 TSLP protein is post-translationally modified by the endogenous protease. The cleaved TSLP shows higher activity, producing IL-5 when stimulated with IL-1β, than the full-length form.40 Of interest, Kim et al.44 authors recently demonstrated that TSLP production was induced by periostin in epithelial cells under the Th2 high inflammatory condition like eosinophilic NP. Until now, TSLP has been consistently reported to play a pathological role in eosinophilic NP unlike IL-25 and IL-33.
ILCs
Epithelial-derived cytokines, such as IL-25, IL-33, and TSLP, exert effects on type 2 ILCs (ILC2s).45 ILCs are lymphocyte-like cells, but lack markers of mature lymphocytes, and do not express allergen-specific T cell receptors. ILC2s are regarded as innate counterparts of Th2 cells because both share the same functional module by their mutual production of signature cytokines, such as IL-5 and IL-13.46 For example, GATA-3 is a key transcriptional factor that plays parallel roles in the development and function of both Th2 cells and ILC2s.47 Moreover, the signal transducer and activator of transcription (STAT)-6 is also an important factor for Th2 polarization and plays a role in the post-developmental role in ILC2s, though it is not required for the development of ILC2s.46 Interestingly, IL-33- and IL-25-activated ILC2s can induce eosinophilic airway inflammation accompanied by airway hyperresponsiveness even in recombination-activating gene (Rag) knockout mice, which means ILC2s have function independent of acquired immunity.4849 ILC2s are abundant and also have a close relationship with higher tissue and blood eosinophilia in NP, clinically related to worsening nasal symptom scores and asthma comorbidity.5051 Bal et al.52 have reported that there is spatial co-localization between ILC2s and eosinophils in NP, and that co-culture of eosinophils and ILC2s augmented the activation of eosinophils and prolonged their survival, and in return pre-activated eosinophils enhanced IL-5 production of ILC2s in an IL-4 dependent manner. Of note, ILC2s have functional plasticity responsive to environmental cues, including viral infection. Mouse ILC2s in the lung undergo T-bet-mediated plasticity in response to infection, including influenza virus, respiratory syncytial virus, Haemophilus influenza, and Staphylococcal aureus.17 Human ILC2s can be converted into ILC1s by IL-12 and reversed by IL-4,52 or into interferon (IFN)-γ/IL-13 dual-producing ILC1s in response to both IL-1β and IL-12.53 T cells that are able to produce both IFN-γ and IL-13 induce enhanced airway hyperresponsiveness compared to conventional Th2 cells.54 Thus, ILC2 plasticity may contribute to disease heterogeneity which might lead to recalcitrance and exacerbation of inflammatory diseases.
T cell subsets
Three major subsets of CD4+ T effectors, classified as Th1, Th2, and Th17, function in host defense in response to various types of pathogens and are involved in different types of tissue injury in immunologic diseases. Regulatory T cells (Tregs) modulate T cell response inducing the termination of inflammation. Definite characteristics of differentiated subsets of T effector cells are cytokines they produce and transcriptional factors they express. Signature cytokines produced by the major CD4+ T cell subsets are IFN-γ for Th1 cells; IL-4, IL-5, and IL-13 for Th2 cells; IL-17 and IL-22 for Th17 cells, and IL-10 and transforming growth factor-beta (TGF-β) for Treg. Involved transcriptional factors are T-bet for Th1, GATA-3 for Th2, retinoid-related orphan receptor C (RORC) for Th17, and FOXP3 for Treg. CRS was initially classified according to the presence of NP which reflects predominantly eosinophil-infiltrating Th2 inflammation at the molecular levels in Western countries.55 However, Asian populations, including Chinese population, showed multiple inflammatory cell infiltrates, including neutrophils and T cells classified as Th1/Th17 cells.56 Furthermore, mixed Th17/Th2 inflammatory patterns were also demonstrated throughout a single nasal polyp tissue, and eosinophilic NP also encompass neutrophils and their biological markers such as IL-8 and myeloperoxidase.2 Therefore, NP begin to be understood from the dichotomization of Th1 or Th2 to a multi-dimensionally evolved concept. A European multicenter case-control study group (GA2LEN Sinusitis Cohort group) recently conducted the clustering of CRS subjects by T cell subset-based cytokines and explored whether these cytokines adequately reflect phenotype, such as the presence of NP or asthma comorbidities.57 This study analyzed tissue cytokines from 173 CRS subjects and 89 controls in a phenotype-free manner and classified 10 clusters: 4 clusters with low or undetectable IL-5, immunoglobulin E (IgE), and eosinophilic cationic protein (ECP); and 6 clusters having moderate to high concentrations of these markers. In detail, cluster 1 has minimal inflammatory markers, cluster 2 associated with Th22, cluster 3 representing Th1, and cluster 4 showing mixed Th17/Th22/Th1. The latter 6 clusters with elevated IL-5 were classified into 2 groups: clusters 8-10 showed the highest IL-5 with enhanced IgE against Staphylococcal enterotoxins plus neutrophilic markers plus Th17/Th22 cytokines implying sophisticated pathomechanism, whereas clusters 5-7 showed mainly IL-5 dominant CRS. This clustering roughly reflects phenotypes: clusters 1-3 represent CRS without NP (CRSsNP) and low asthma comorbidity, but the proportion of NP and asthma comorbidities seems to increase from cluster 4 to cluster 10. Furthermore, it was reported to be a distinct regional difference in endotypes.2 CRSwNP tissues from patients of European countries, such as Benelux and Berlin, had a high proportion of IL-5 (62%-82%); those from Adelaide, Australia showed IL-5 in 46% and mixed IL-5 plus IL-17 or IFN-r in 27%. Most of the Western NP were Th2-biased. However, there was a remarkable difference even in Chinese NP. Beijing NP showed mixed IL-5/IL-17 portion of 42%; NP in Chengdu, a Chinese city, mainly had a negative cytokine pattern of 57%. These results imply that CRS is not a dichotomous disease but has multi-dimensional continuum worldwide. To make this concept clearer and more understandable, longitudinal cohort studies would be required rather than cross-sectional, case-control studies conducted in the past.
Remodeling
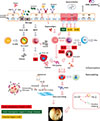
CRS is characterized by edematous remodeling patterns, basement membrane thickening, and goblet cell hyperplasia.5859 TGF-β is not only a chemoattractant for fibroblasts, enhancing their proliferation and collagen deposition, but also an inducer for Treg. High TGF-β is characteristic for CRSsNP, but low TGF-β was a signature finding in patients with CRSwNP.5860 Low TGF-β is also associated with a defect of Treg, namely, failing to maintain homeostasis, additionally, failure of deposition of collagens and other ECMs. High matrix metallopeptidases (MMPs), which are usually observed in NP, support ECM degradation.61 Besides the roles of TGF-β in the remodeling process in NP, the dysregulated coagulation system can be engaged in this process. Coagulation and fibrinolysis cascades are the biological processes involved in fibrin deposition and dissolution.62 The first study to suggest local activation of the coagulation system in sinus tissue showed that the concentration of thrombin and thrombin-antithrombin complex were increased in CRSwNP, which in turn participated in the remodeling process via a vascular endothelial growth factor pathway.63 Since that study, well-conducted reports have documented that the down-regulation of tissue plasminogen activator (t-PA) and activated coagulation factor XIIIa are involved in fibrin deposition and enhancement of fibrin crosslinking, culminating in edematous remodeling patterns in NP.636465 Of interest, the concentration of t-PA in CRS tissues shows a negative correlation with the concentration of ECP, a marker of eosinophils, and t-PA is mainly stained in epithelial cells on immunohistochemistry and down-regulated in epithelial cells by Th2 cytokines, such as IL-4 and IL-13.64 Furthermore, M2 macrophages, alternatively activated by Th2 cytokines,66 produce coagulation factor XIIIa which acts enzymatically and contribute to the formation of a tight tetrameric complex (FXIIIA2B2), a cross-linking process of fibrin.65 In summary, Th2-induced coagulopathy causes excessive fibrin deposition to induce edematous remodeling patterns in NP. The pathogenesis of NP described above is depicted in Figure.
PERSONALIZED THERAPEUTIC STRATEGIES
Therapeutic strategies according to clinical or pathologic markers have also been developed. Among these signs, tissue eosinophilia is the most important regarding disease recurrence and comorbidities.7 Approximately 70%-80% of CRSwNP cases accounts for eosinophilic types in the Western country, whereas 30%-40% cases constitute eosinophilic NP in Asian countries, including South Korea and China.29 Eosinophilic CRS is a better responder to steroid therapy vs non-eosinophilic CRS.67 Non-eosinophilic CRS seems to be a better responder to surgical treatment vs eosinophilic CRS, especially in elderly subjects.68 Therefore, how to discriminate between 2 groups by using clinical features or easily available samples, such as blood samples, has been investigated because tissue eosinophil count using biopsy tissue is not practical for primary care clinicians. Blood eosinophilia, computerized tomography (CT) scans showing ethmoidal dominance, bilateral disease, asthma comorbidity, and aspirin sensitivity are predictors of eosinophilic NP.6970 Despite strategical approaches according to histologic or clinical subtypes, there must be mixed pathologic subgroups in recalcitrant subgroups which require further innovative therapeutic plans. That is why endotyping and the development of biologicals corresponding to their endotypes are inevitable at present. When we determine molecular targets from previous research data, Th2 cytokines, such as IL-4, IL-5, IL-13, and periostin, epithelial-derived cytokines, such as TSLP, IL-33, and IL-25, IgE as an acquired immunity marker, and Th17 cytokines such as IL-17a and IL-22, can be candidates for biologicals. Among these, several human monoclonal antibodies became commercially available or under clinical trials in allergic disease.
A humanized monoclonal anti-IgE antibody, omalizumab, which binds to free IgE with high affinity, has been used in moderate to severe allergic asthma in the US and Europe.71 Hanania et al.72 have demonstrated that periostin is a marker for omalizumab responsiveness in patients with moderate to severe allergic asthma. Since the concentration of IgE is locally increased in eosinophilic NP,73 a strategy for antagonizing IgE might be relevant in subjects with eosinophilic NP. However, a clear indication of omalizumab remains undetermined in NP.74 Given that periostin is upregulated and secreted from mast cells via IgE-mediated signaling in eosinophilic NP,44 periostin might be a biomarker for responsiveness to omalizumab in eosinophilic NP. Gevaert et al.75 conducted a randomized, double-blind, placebo-controlled study on the effects of omalizumab on NP and comorbid asthma (n=24). A significant decrease in the total nasal endoscopic polyp score after 16 weeks in the omalizumab-treated group was observed. Omalizumab also showed significant benefits in nasal and respiratory symptoms, such as nasal congestion, anterior rhinorrhea, loss of smelling sense, wheezing, and dyspnea, as well as quality-of-life scores, irrespective of the presence of allergy. However, there was no reduction in nasal or serum inflammatory mediators in the treated group. Larger sizes of clinical trials are needed to confirm this proof-of-concept study and to analyze subgroups for appropriate biomarkers to guide better treatment responses.
Eosinophils, mast cells, and ILC2s mainly produce IL-5 which is a strong driver to Th2 inflammation and associated with a higher risk of having asthma comorbidity.76 As mentioned above, 80% of Western NP cases account for the eosinophilic type. Furthermore, over 80% of European NP cases and 20%-60% of Asian NP cases expressed IL-5 in NP tissue homogenates.2 Based on previous mechanistic studies, antagonizing IL-5 would be considered a good therapeutic target. In a small clinical trial, subjects treated with anti-IL-5 mAb (reslizumab) showed reduced polyp size, blood eosinophilic counts, and ECP concentration in nasal secretions. Responders had higher IL-5 levels in nasal secretions compared with non-responders. However, there was no symptom improvement in the treated group.77 A clinical trial of another anti-IL-5 mAb, mepolizumab, demonstrated similar results as a previous study and showed a significant reduction in polyp score, blood eosinophilic count, serum ECP, and serum IL-5Rα, but no significant improvement in symptom scores.78 based on earlier studies, it still remains unclear which biomarker can be used to select good responders to anti-IL-5 treatment.
Dupilumab targeting IL-4Rα has inhibitory effects on the signaling of IL-4 and IL-13 since both cytokines signaled through IL-4Rα as a common subunit. IL-4 and IL-13 act through 2 different receptors. One is type 1, activated by IL-4 only and expressed on the lymphocyte. The other is type 2 receptor activated by both IL-4 and IL-13 and expressed by various cells.71 Blocking the type 2 receptor is promising for controlling recalcitrant allergic disease.7980 Since type 2 receptors can play a pivotal role in polypogenesis, dupulimab may play a role in treating recalcitrant NP. Recently, a multi-center clinical trial demonstrated a potential role of dupilumab in NP showing a significant decrease in nasal polyp score and improvements in olfaction and CT scores as well as other clinical outcomes, such as nasal symptoms and quality of life.81
Epithelial cell-derived innate cytokines, including TSLP, exert major effects on Th2 inflammation. However, there has been no study regarding its effect on NP, although a previous study investigated the inhibitory effects of anti-TSLP on allergen-induced asthmatic response.82 The roles of IL-25 and IL-33 in NP remain unclear and need further investigation.83 Therefore, targeting epithelial derived innate cytokines can be promising because it may control upstream mediators which T cell subsets do not act on.
CONCLUSIONS
A clinically observable phenotype includes multiple molecular endotypes with different prognoses. Therefore, phenotyping is not sufficient to predict responsiveness to medical or surgical treatments and the risk of comorbid conditions. With the advent of an era with biologicals, endotyping helps select patients suitable for each biological which can be a breakthrough in treating NP. Although modulating acquired immunity, for example T cell subsets, has made some progression, targeting epithelial cell-derived innate cytokines, such as TSLP, IL-33, and IL-25, may provide novel opportunities to manage CRS and NP as well as allergic airway disease.
 XML Download
XML Download